XiaoMi-AI文件搜索系统
World File Search Systemtransduced

基于 CRISPR/Cas9 的银叶粉虱 (Bemisia tabaci) 基因组编辑
烟粉虱隐种中东-小亚细亚 I (MEAM1) 是一种严重的农业广食性害虫,也是多种植物病毒的载体,在全球范围内造成了巨大的经济损失。由于缺乏强大的基因编辑工具,烟粉虱的控制受到限制。烟粉虱的基因编辑很困难,因为其胚胎很小,在技术上很难注射,而且注射后死亡率很高。我们开发了一种 CRISPR/Cas9 基因编辑方案,该方案基于注射卵黄发生成年雌性而不是胚胎(“ReMOT 控制”)。我们鉴定了一种卵巢靶向肽配体(“BtKV”),当它与 Cas9 融合并注射到成年雌性体内时,会将核糖核蛋白复合物转导至生殖系,从而实现对后代基因组的有效、可遗传的编辑。与胚胎注射相比,成虫注射很容易,并且不需要专门的设备。开发易于使用的烟粉虱基因编辑协议将使研究人员能够将反向遗传方法应用于该物种,并将带来针对这种毁灭性害虫的新控制方法。

与基因工程T细胞的免疫疗法有望治疗狗的非恶性疾病
T细胞激活所需的。使用慢病毒或复古病毒载体将所得的汽车结构永久整合到患者T细胞的基因组中,从而导致汽车在转导的T细胞上永久表达。然后将细胞送回患者(自体疗法) - 降低淋巴结障碍的预处理方案(图2)。CAR -T细胞具有其抗原特异性 - 有效地重定向,现在通过SCFV以主要的兼容性非依赖性方式通过SCFV实现了抗原识别。在与靶细胞表面表达的Cognate抗原接合后,CAR -T细胞会经历抗Gen特异性激活,增殖,细胞因子促进和细胞毒性。现在已经证明了使用慢病毒和逆转录病毒技术生成犬T细胞的可行性,并证实了它们在体外经历抗原特异性T细胞激活和效应子功能的能力。2–6个优化的制造方案现已发布针对犬T细胞,在几个重要方面与人类CAR-T细胞的制造有所不同,包括
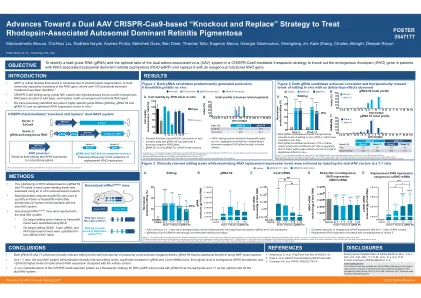
基于双 AAV CRISPR-Cas9 的“...
mRho hRHO/+ 小鼠注射了双 AAV 系统,其中以不同的载体 1:载体 2 比例识别出领先的 gRNA 59,并在注射后 6 周进行分析(载体 n=12;gRNA 59 n=20–22)。显示平均值 (SD)。*p<0.05、**p<0.01、***p<0.001、****p<0.0001 vs 载体。# p<0.05、## p<0.01、#### p<0.0001 vs 其它载体比例。(A) 编辑标准化为转导区域。黑色虚线表示达到治疗相关编辑水平 (≥25%) 的阈值。3 (B) gRNA 水平。(C) Cas9 mRNA 水平。(D) 内源性 hRHO mRNA 水平。数据标签表示与载体相比的百分比下降。(E) 外源性替代 coRHO mRNA 水平。数据标签表示与载体相比的倍数增加。AAV,腺相关病毒;bp,碱基对;coRHO,密码子优化的RHO等位基因;gRNA,向导RNA;hRHO,人类RHO等位基因;mRho,小鼠Rho等位基因;NGS,下一代测序;RHO/Rho,视紫红质;SD,标准差。
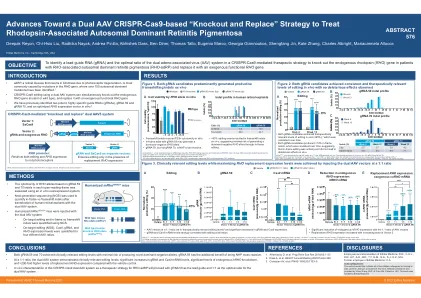
朝着基于双AAV CRISPR-CAS9的双重前进
MRHO HRHO/+小鼠在各种矢量1:矢量2比率下注射了双AAV系统,并在注射后6周分析了铅GRNA 59,并在注射后6周进行分析(车辆n = 12; n = 20-22的GRNA 59)。平均(SD)。*p <0.05,** p <0.01,*** p <0.001,**** p <0.0001与车辆。#p <0.05,## p <0.01,#### p <0.0001 vs其他向量比。(a)编辑归一化为转导面积的编辑。黑色虚线表示实现治疗相关的编辑水平(≥25%)的阈值。3(b)GRNA水平。(c)Cas9 mRNA水平。(d)内源性HRHO mRNA水平。数据标签表明%降低与车辆。(E)外源替代Corho mRNA水平。数据标签表明折叠与车辆增加。AAV,腺相关病毒; BP,基对; Corho,密码子优化了Rho等位基因; GRNA,导向RNA; Hrho,人类Rho等位基因; Mrho,老鼠Rho等位基因; NGS,下一代测序; Rho/Rho,Rhodopsin; SD,标准偏差。
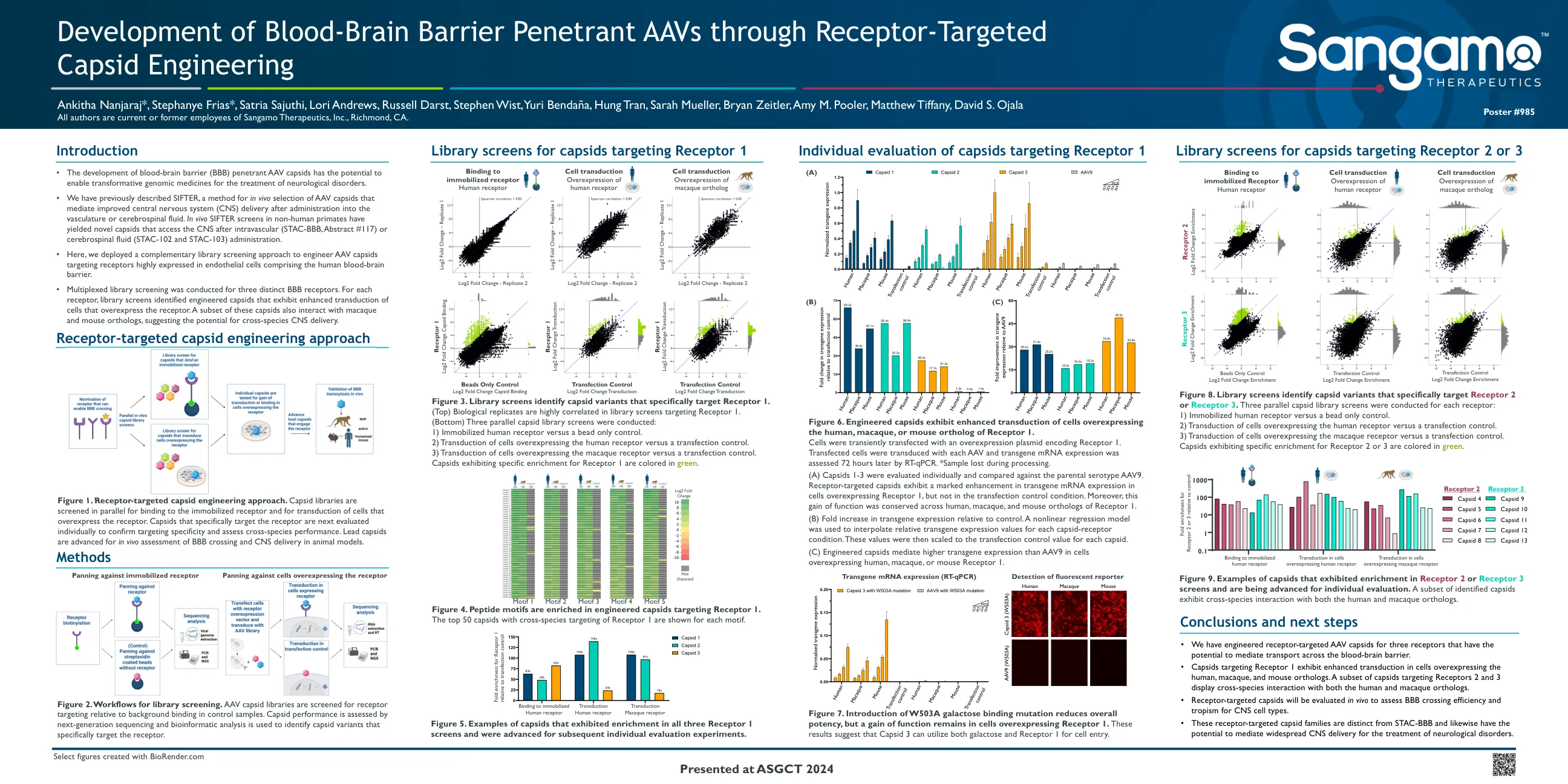
靶向受体的衣壳工程方法...
图6。工程的衣壳表现出增强的过表达人,猕猴或小鼠直系同源物的细胞的转导增强。细胞用过表达质粒编码受体1。转染的细胞用每种AAV转导,并在72小时后通过RT-QPCR评估转基因mRNA表达。*处理过程中丢失的样本。(a)CAPSIDS 1-3单独评估并与父母血清型AAV9进行比较。受体靶向的衣壳在过表达受体1的细胞中表现出明显的转基因mRNA表达,但在转染控制条件下却没有明显增强。此外,这种功能的增益在受体的人,猕猴和小鼠直系同源物中得到了保守。(b)相对于对照的转基因表达的倍数增加。一个非线性回归模型用于每个衣壳受体条件的相对转基因表达值。然后将这些值缩放到每个衣壳的转染控制值。(C)在过表达人,猕猴或小鼠受体1的细胞中,工程上的衣壳介导更高的转基因表达。

直接注射的慢病毒载体 T 细胞疫苗可保护小鼠免受急性和慢性病毒感染
基于慢病毒载体的树突状细胞疫苗在动物模型中诱导保护性 T 细胞反应,以抵抗病毒感染和癌症。在本研究中,我们测试了是否可以通过直接注射表达抗原的慢病毒载体来实现预防性和治疗性疫苗接种,从而避免体外转导树突状细胞。注射的慢病毒载体优先转导脾脏树突状细胞并导致长期表达。注射编码淋巴细胞脉络丛脑膜炎病毒 (LCMV) 的 MHC I 类限制性 T 细胞表位和 CD40 配体的慢病毒载体可诱导抗原特异性细胞溶解性 CD8 + T 淋巴细胞反应,从而保护小鼠免受感染。向慢性感染小鼠注射编码 LCMV MHC I 类和 II 类 T 细胞表位和可溶性程序性细胞死亡 1 微体的慢病毒载体可迅速清除病毒。通过直接注射慢病毒载体进行疫苗接种对无菌 α 基序和含有 HD 结构域的蛋白 1 敲除 (SAMHD1 敲除) 小鼠更有效,这表明含有 Vpx(一种通过诱导 SAMHD1 降解来提高树突状细胞转导效率的慢病毒蛋白)的慢病毒载体将成为治疗人类慢性疾病的有效策略。

直接注射的慢病毒载体 T 细胞疫苗可保护小鼠免受急性和慢性病毒感染
基于慢病毒载体的树突状细胞疫苗在动物模型中诱导保护性 T 细胞反应,以抵抗病毒感染和癌症。在本研究中,我们测试了是否可以通过直接注射表达抗原的慢病毒载体来实现预防性和治疗性疫苗接种,从而避免体外转导树突状细胞。注射的慢病毒载体优先转导脾脏树突状细胞并导致长期表达。注射编码淋巴细胞脉络丛脑膜炎病毒 (LCMV) 的 MHC I 类限制性 T 细胞表位和 CD40 配体的慢病毒载体可诱导抗原特异性细胞溶解性 CD8 + T 淋巴细胞反应,从而保护小鼠免受感染。向慢性感染小鼠注射编码 LCMV MHC I 类和 II 类 T 细胞表位和可溶性程序性细胞死亡 1 微体的慢病毒载体可迅速清除病毒。通过直接注射慢病毒载体进行疫苗接种对无菌 α 基序和含有 HD 结构域的蛋白 1 敲除 (SAMHD1 敲除) 小鼠更有效,这表明含有 Vpx(一种通过诱导 SAMHD1 降解来提高树突状细胞转导效率的慢病毒蛋白)的慢病毒载体将成为治疗人类慢性疾病的有效策略。

系统性外泌体导入后淋巴细胞的体内工程...
使用逆转录病毒或慢病毒载体转导的干细胞或 T 细胞进行体外基因治疗,在治疗免疫缺陷和癌症方面已显示出显著的疗效。然而,这个过程成本高昂,技术难度大,而且不易推广到大量患者群体,特别是在世界欠发达地区。直接体内基因治疗可以避免这些问题,而且在临床试验中,腺相关病毒 (AAV) 载体的这种方法已被证明对影响肝脏和中枢神经系统等分化组织的疾病是安全有效的。然而,在全身给药后用 AAV 在体内转导淋巴细胞的能力尚未得到仔细探索。在这里,我们表明,在小鼠全身给药后,AAV8 载体的标准制剂和外泌体相关制剂都可以有效转导各种免疫细胞群,包括 CD4 + T 细胞、CD8 + T 细胞、B 细胞、巨噬细胞和树突状细胞。我们通过检测 AAV 基因组和转基因 mRNA 提供了 T 细胞转导的直接证据,并表明可以表达细胞内和跨膜蛋白。这些发现确立了 AAV 介导的体内基因递送至免疫细胞的可行性,这将促进基础研究和应用研究,以实现直接体内基因免疫疗法的目标。

CRISPR 筛选可识别调控黑色素瘤细胞侵袭性的基因
恶性癌细胞会不受控制地增殖,并可能转移到远处器官。转移的一个关键步骤是癌细胞在扩散到远处器官之前侵入邻近组织的能力。因此,了解侵袭机制可能有助于发现新的可用药物靶点以防止转移。在本项目中,使用 CRISPR 敲除筛选体外研究了人类黑色素瘤细胞的侵袭性。为此,评估了三个汇集的 CRISPR 文库,然后选择其中一个进行筛选。在验证了所选表观遗传敲除文库的 gRNA 表示后,生成了一个慢病毒文库以转导 A375 黑色素瘤细胞。然后使用 Matrigel 侵袭室通过优化的侵袭试验检查突变黑色素瘤细胞的侵袭性。将转导的黑色素瘤细胞接种到上室中,并使其通过 Matrigel 迁移到具有更高化学引诱剂浓度的下室。随后分别收集上室和下室细胞,分离基因组DNA,通过PCR扩增制备测序文库,并用Illumina新一代测序技术进行测序。本报告不包含CRISPR筛选的测序数据。

Invitrogen™ LentiArray™ 人类 CRISPR 文库,96 孔...
转导条件 • 您必须根据经验确定每种细胞系的转导条件和感染复数 (MOI)。如果需要共感染,我们建议对 Cas9 至 CRISPR 文库慢病毒颗粒使用 5–10 的 MOI 比率,以达到最佳基因敲除程度。 • 感染期间使用含较低水平 FBS(例如 3– 5% FBS)的培养基可能会增加某些细胞类型的转导效率。 • Polybrene™(六甲溴铵)可以将慢病毒转导到人细胞的效率提高 2–10 倍。您必须根据经验确定目标细胞的最佳 Polybrene™ 浓度(例如最大感染性,最小毒性)。我们建议使用浓度范围(2–8 µg/mL)对 Polybrene™ 耐受性进行初步测试。 • 如果您计划使用嘌呤霉素进行选择,则必须首先确定选择转导细胞所需的最佳嘌呤霉素浓度。抗生素批次、细胞类型、细胞生长动力学和细胞培养条件(包括细胞密度)会影响筛选所需的嘌呤霉素量。使用嘌呤霉素进行筛选通常需要 7-10 天。