XiaoMi-AI文件搜索系统
World File Search Systemtransgenic

利用靶向捕获测序鉴定转基因作物中的T-DNA结构和插入位点
转基因作物的商业化需要严格的安全评估,包括对插入的 T-DNA 进行精确的 DNA 水平表征。过去,已经开发了几种识别 T-DNA 插入位点的策略,包括南方印迹和不同的基于 PCR 的方法。然而,这些方法通常难以扩大规模以筛选数十种转基因事件和具有复杂基因组的作物,如马铃薯。在这里,我们报告使用目标捕获测序 (TCS) 来表征马铃薯中 34 个转基因事件的 T-DNA 结构和插入位点。这个 T-DNA 是左右边界之间的 18 kb 片段,携带三个抗性 (R) 基因(RB、Rpi-blb2 和 Rpi-vnt1.1 基因),可完全抵抗晚疫病。使用 TCS,我们在 T-DNA 和连接区域内获得了高序列读取覆盖率。我们确定了 85% 转基因事件两端的 T-DNA 断点。约 74% 的转基因事件的 T-DNA 中 3 个 R 基因序列完整。一半转基因事件的 T-DNA 侧翼序列来自马铃薯基因组,约三分之一 (11) 的转基因事件在马铃薯基因组中定位了一个 T-DNA 插入,其中五个事件不会中断现有的马铃薯基因。使用 PCR 和 Sanger 测序确认了 6 个最佳转基因事件的 TCS 结果,这 6 个转基因事件占适合监管部门批准的转基因事件的 20%。这些结果证明了 TCS 在转基因作物中精确表征 T-DNA 插入方面具有广泛的适用性。

提高非生物胁迫耐受性的转基因育种方法:最新进展和未来展望
摘要:近年来,全球气候变化迅速,人口不断增加,导致非生物胁迫发生率增加,农作物产量下降。环境胁迫,如温度、干旱、营养缺乏、盐度和重金属胁迫,是农业面临的主要挑战,它们导致农作物生长和产量大幅下降。非生物胁迫是一种非常复杂的现象,涉及植物细胞的各种生理和生化变化。暴露于非生物胁迫的植物表现出活性氧 (ROS) 水平的提高,这种物质具有高活性和毒性,会影响叶绿素的生物合成、光合能力以及碳水化合物、蛋白质、脂质和抗氧化酶的活性。转基因育种为实现植物遗传改良提供了一种合适的常规育种替代方案。在过去的二十年里,基因工程/转基因育种技术在操纵基因以诱导转基因植物所需特性方面取得了显著进展。转基因方法使我们能够识别参与特定植物过程的候选基因、miRNA 和转录因子 (TF),从而能够全面了解影响植物耐受性和生产力的分子和生理机制。这种现象的准确性和精确性确保了未来植物改良的巨大成功。因此,转基因育种已被证明是改善作物非生物胁迫的一种有前途的工具。本综述重点介绍了转基因育种在提高植物非生物胁迫耐受性和生产力方面的潜在和成功应用、最新进展和未来前景。

一套增强子 AAV 和转基因小鼠系,用于 1 基因访问皮质细胞类型 2
Yoav Ben-Simon, 1,4 Marcus Hooper, 1,4 Sujatha Narayan, 1,4 Tanya Daigle, 1,4 Deepanjali Dwivedi, 1 Sharon W. 4 Way, 1 Aaron Oster, 1 David A. Stafford, 2 John K. Mich, 1 Michael J. Taormina, 1 A. Refugio, 1 A. Martina-Jamena. R. Roth, 1 Shona Allen, 2 Angela Ayala, 1 Trygve E. Bakken, 1 Tyler Barcelli, 1 Stuard Barta, 1 6 Jacqueline Bendrick, 1 Darren Bertagnolli, 1 Jessica Bowlus, 1 Gabriella Boyer, 1 Krissy Brouner, 1 Brittny Casian, 1 7 Chara Chair, Chara Rush, 1 Chara Rush. barty, 1 Rebecca K. Chance, 2 Sakshi Chavan, 1 Maxwell 8 Departee, 1 Nicholas Donadio, 1 Nadezhda Dotson, 1 Tom Egdorf, 1 Mariano Gabitto, 1 Jazmin Garcia, 1 Amanda 9 Gary, 1 Molly Gasperini, 1 Jeffry Goldy, 1 1 Blanche, 1 Lucas Gregory, No. . 1 Francoise Haeseleer, 1 10 Carliana Halterman, 1 Olivia Helback, 1 Dirk Hockemeyer, 2 Cindy Huang, 1 Sydney Huff, 1 Avery Hunker, 1 Nelson 11 Johansen, 1 Zoe Juneau, 1 Brian Kalmbach, 1 Shannon Khem, 1 Emily Kuckel, 1 Lar Rasen, 1 12 Changkyu Lee, 1 Angus Y. Lee, 2 Madison Leibly, 1 Garreck H. Lenz, 1 Elizabeth Liang, 1 Nicholas Lusk, 1 Jocelin 13 Malone, 1 Tyler Mollenkopf, 1 Elyse Morin, 1 Dakota Newman, 1 Lydia Ng, 1 Kiet Ngoste, 1 1 Victoria Oman, 14 h Pham, 1 Christina A. Pom, 1 Lydia Potekhina, 1 Shea Ransford, 1 Dean Rette, 1 Christine 15 Rimorin, 1 Dana Rocha, 1 Augustin Ruiz, 1 Raymond EA Sanchez, 1 Adriana Sedeno-Cortes, 1 Joshua P. Sevigny, 1 Nadi Lava, 16 Lyvalomi Ana R. Sigler, 1 La' Akea Siverts, 1 Saroja Somasundaram, 1 Kaiya 17 Stewart, 1 Eric Szelenyi, 1 Michael Tieu, 1 Cameron Trader, 1 Cindy TJ van Velthoven, 1 Miranda Walker, 1 Natalie 18 Weed, 1 Morgan Wirlin, 1 Toren Wood, 1 Toren Wood, 1 Zilda o, 1 Thomas Zhou, 1 Jeanelle Ariza, 1 Nick 19 Dee, 1 Melissa Reding, 1 Kara Ronellenfitch, 1 Shoaib Mufti, 1 Susan M. Sunkin, 1 Kimberly A. Smith, 1 Luke 20 Esposito, 1 Jack Waters, 1 Bargavi Thyagarajan, 1 Yaqin , 1 Shenq , 1 Sheng Leng . Boaz P. Levi, 1 John 21 Listen, 2,3 Jonathan Ting, 1 Bosiljka Tasic 1,5,* 22

利用 CRISPR/Cas9 技术生成和鉴定转基因人类多能干细胞的流程
由于其无限的增殖潜力、整倍体状态以及向任何细胞类型分化的能力,人类多能干细胞 (hPSC)(无论是胚胎细胞还是诱导细胞)在疾病建模和生产临床应用细胞方面具有巨大潜力 [ 1 – 3 ]。尽管已经建立了来自患有各种疾病的患者的许多 hPSC 系,但是针对某些病理或罕见基因突变生成 hPSC 系仍然具有挑战性。此外,个体间的遗传异质性可能导致生物学变异,从而使系间比较困难,尤其是来自健康对照和患者的 hPSC 之间的比较 [ 4 , 5 ]。对 hPSC 进行遗传操作的能力为我们引入、修改或校正突变以及生成遗传匹配的同基因对照系提供了机会,从而建立明确的基因型-表型关联 [ 6 , 7 ]。近年来,基于位点特异性核酸酶(包括锌指核酸酶 (ZFN)、转录激活因子样效应核酸酶 (TALEN),尤其是成簇的规律间隔短回文重复序列 (CRISPR) 系统)的技术已使 hPSC 的基因组工程变得十分灵活 [8,9]。然而,由于 hPSC 的固有特性,包括相对较差的转染效率和转染后存活率低、难以分离克隆群、优先选择和扩增非整倍体克隆以及自发细胞分化,hPSC 工程仍然具有挑战性。为了缓解这些问题,已经描述了几种用于产生各种不同诱变事件的方案 [10-14]。尽管人们投入了大量精力来改进产生转基因 hPSC 的方法程序,但只有少数研究

动物中的重组或合成DNA使用:
• Creation of transgenic or knock-out animals, other than rodents, made by stable introduction of foreign DNA • Breeding of transgenic or knock-out animals, other than rodents, made by stable introduction of foreign DNA • Purchase or transfer of transgenic or knock-out animals, other than rodents, made by stable introduction of foreign DNA • Experiments in which rDNA molecules are administered to a transgenic animal • Experiments in which viral vectors containing将rDNA分子施用对任何动物(转基因或其他方式)•将通过rDNA分子修饰的细胞(包括从转基因动物分离的细胞)施用的实验对任何动物(转基因或否则)施用•通过RDNA分子修饰的微生物对任何动物(否则)(否则)否则(否则)否(否则)

通过转基因阵列在秀丽隐杆线虫中的高通量文库转基因导致综合序列多样性(TARDIS)
图5。TARDIS启动子库。a)概述两个分裂的着陆垫及其相关的启动子插入向量。正确整合后,选择性标记和荧光团表达都会恢复。b)从单个TARDIS阵列线的单个热轴(PX819)中回收了9个基因的转录记者。集成到单个McSarlet-I /Hygr着陆垫中。主图像显示了指定的报告基因的MSCARLET-I表达,而插图显示同一区域的极化图像。c)示例同时,从单个TARDIS阵列中的双重整合到带有害虫的双降落垫菌株中。ceh-10p :: mneongreen :: pest是假彩色绿色和ceh-40p :: mcarlet-i :: pest是假彩色的洋红色。所有比例尺均代表20µm

交联的肌动蛋白网络(氏族)会影响转基因转化和原代人小梁网络中的刚度和/或活性动力学
小梁网(TM)细胞中的交联肌动蛋白网络(氏族)可能通过改变TM细胞功能和刚度来增加IOP。但是,缺乏直接证据。在这里,我们开发了转化的TM细胞,形成自发荧光标记的氏族。通过将转化的青光眼TM(GTM3)细胞与柔抗脱反应-EGFP-BLASTR慢病毒载体载体并用BlastCidin选择,构建了稳定的细胞。使用原子力显微镜研究了GTM3-氟法中GFP细胞的刚度。还测量了用/不含地塞米松/TGFβ2处理的原代人TM细胞中氏族的弹性模量,以验证在GTM3-氟法中GFP细胞中的发现。对用1μM拉氏蛋白B或Phrodo Bioparticle处理的GTM3-氟法中的活细胞成像分别确定肌动蛋白稳定性和吞噬作用。GTM3-脱反性GFP细胞形成自发氏族,而无需诱导TGFβ2或地塞米松。与没有氏族的细胞相比,含有细胞的氏族显示出升高的细胞刚度,对latrunculin b诱导的肌动蛋白去聚合的抗性以及造成的吞噬作用。用来塞米松或TGFβ2诱导的氏族的原代人TM细胞也被僵硬,吞噬细胞较少。GTM3- LIFEACT-GFP细胞是研究TM中氏族的机械生物学和病理学的新工具。这些细胞的初始表征表明,氏族至少有助于TM细胞的一些青光眼表型。
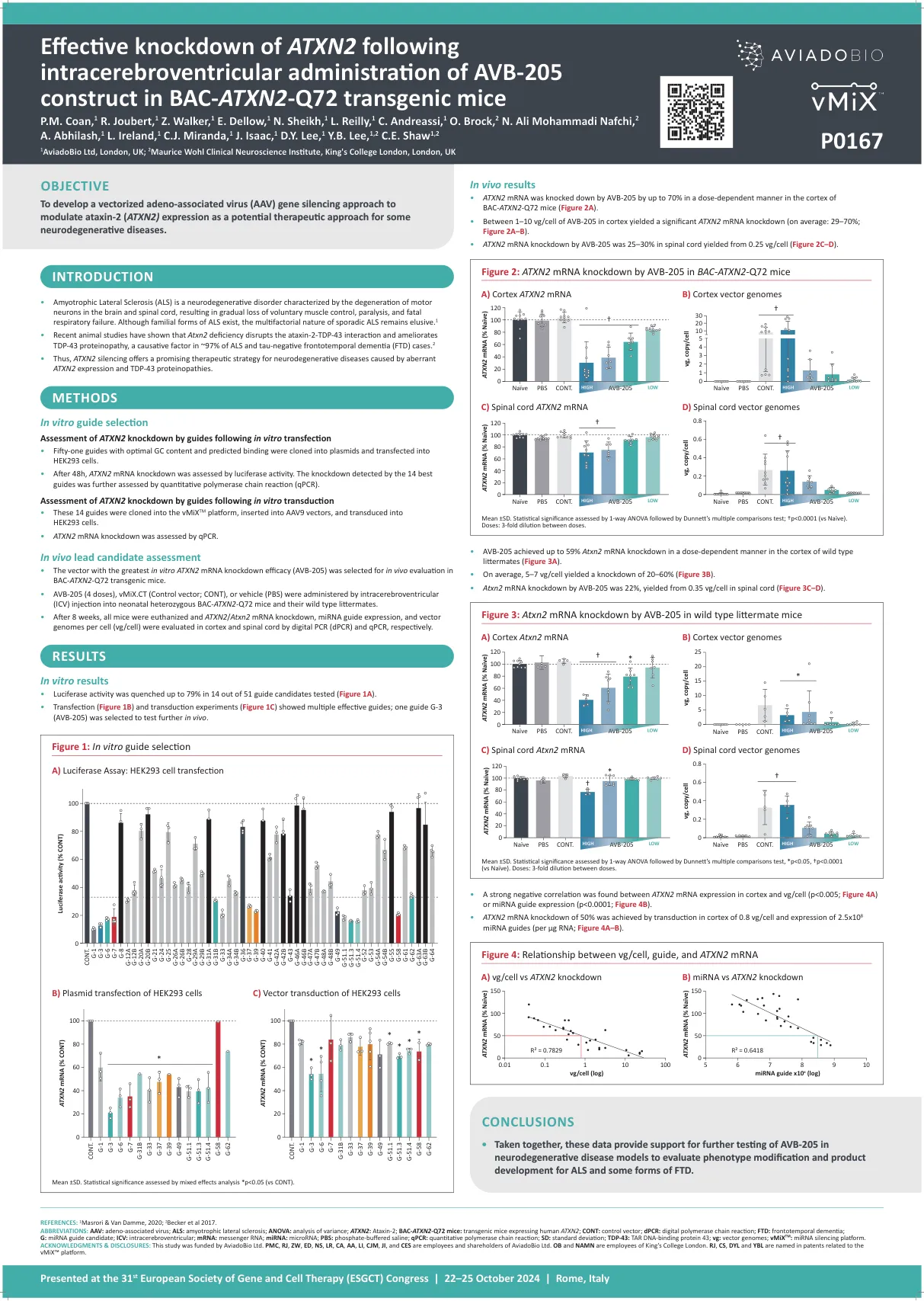
在BACN2 -Q72转基因小鼠中,在脑室内给药后,ATXN2的有效敲低ATXN2
参考文献:1 Masrori&van Damme,2020年; 2 Becker等人2017年。缩写:AAV:腺相关病毒; ALS:肌萎缩性侧索硬化;方差分析:方差分析; ATXN2:ataxin-2; BAC -ATXN2 -Q72小鼠:表达人ATXN2的转基因小鼠;续:控制向量; DPCR:数字聚合酶链反应; FTD:额颞痴呆; G:mirna指南候选人; ICV:脑室室内; mRNA:Messenger RNA; mirna:microRNA; PBS:磷酸盐缓冲盐水; QPCR:定量聚合酶链反应; SD:标准偏差; TDP-43:焦油DNA结合蛋白43; VG:矢量基因组; VMIX TM:miRNA沉默平台。致谢和披露:这项研究由Aviadobio Ltd. PMC,RJ,ZW,ED,NS,LR,CA,CA,AA,LI,CJM,JI和CES资助,是Aviadobio Ltd. OB和NAMN的雇员和股东。RJ,CS,DYL和YBL在与VMIX™平台有关的专利中命名。

分析城市能源以实现净零能源社区
在转基因作物中表达的外源蛋白的细胞定位不仅决定了其稳定性,而且还决定了它们对作物生长和发育的影响,包括在压力条件下;然而,潜在的分子机制仍然未知。在这里,我们通过亚细胞定位,免疫组织化学,免疫流畅和蛋白质印迹分析确定了抗昆虫的转基因水稻huahui-1(HH1)细胞中外源表达的Cry1Ab/c蛋白的细胞分布。通过酵母两杂交,双分子分子荧光互补(BIFC)和辅助药物分析研究了CRY1AB/C蛋白与初筛选的内源性质膜Ca 2+ ATPase之间的相互作用。通过比较CRY1AB/C和Ca 2+ ATPase之间的细胞定位和相互作用位点分析了潜在的相互作用机制。表型指数和Ca 2+ -ATPase活性在转基因HH1和父母线Minghui-63在无压力和盐压力的条件下确定,可以由CRY1AB/C-Ca 2+ -ATPase相互作用调节。结果表明,Cry1ab/C不仅分布在细胞质和核中,而且还分布在质膜上,在质膜上与质膜Ca 2+ -ATPase相互作用。通过BIFC实验,这种相互作用部分保留了细胞核中的质膜蛋白Ca 2+ ATPase,因此可能会通过改变蛋白质的细胞位置来影响膜上Ca 2+ -ATPase活性。一致地,我们的结果证实了转基因HH1中Cry1ab/c的存在导致Ca 2+ -ATPase活性的降低,并对植物表型造成不利影响,包括显着降低的植物高度和生物量,与亲属MH63相比;并且这些有害作用在盐应力条件下更明显,从而影响转基因

使用PiggyBac高效的转基因小鼠产生及其在创始人生成的快速表型
预印本(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。该版本的版权持有人于2023年12月10日发布。 https://doi.org/10.1101/2023.12.10.570953 doi:Biorxiv Preprint