XiaoMi-AI文件搜索系统
World File Search Systemtriggers

据称 YSRCP MP 的裸体视频在网上疯传,引发...
印度民航国务部长 VK Singh 将军表示,随着 Orvakal(Kurnool)新绿地机场的建设和安得拉邦 5 座现有机场的升级,这些机场的总客运处理能力将达到每年约 1000 万名乘客(MPPA)。因此,预计约有 1000 万名乘客将从这些机场受益。在 Lok Sabha 对来自 Kadapa 的 YSRCP MP YS Avinash Reddy 的书面答复中,联邦部长表示,机场的升级/现代化是一个持续的过程,由印度机场管理局 (AAI) 和其他机场运营商不时进行,具体取决于土地的可用性、商业可行性、社会经济因素、航空公司往返此类机场的交通需求/意愿等。在过去五年中,一个绿地机场

基于聚合物PEG的生物正交触发器,用于乳腺癌的前药激活
前药或可以激活前药的成分,特定于肿瘤。生物正交化学已成为按需前药激活的一种有希望的平台,因为它包括可以在生理条件下进行的化学反应而不会干扰生物学过程。4,5这些反应的选择性,特定城市和相当快的动力学允许精确控制非毒性前药的激活。6 - 8据报道,许多生物正交反应具有很高的选择性前药激活的潜力,例如叠氮化物和三苯基芬丁基之间的Staudinger连接,9和跨环环烯(TCO)和四嗪(TZ)之间的四津连接。10,Staudinger连接主要用于连接应用,因为其动力学相对较慢(K 2〜10-3 m-1 s-1),并且少量报告揭示了其前药激活的潜力。11 - 13在低浓度下,四嗪连接以其快速点击释放反应动力学(K 2〜10 4 m-1 s-1)而闻名,许多报告表明,TZ部分的反应性,

通过促进JA信号和Ros-Homeostasis
但是,细胞膜脂质在植物免疫中的作用尚不清楚。我们发现细胞膜定位的磷脂LPE促进了与防御相关的基因表达,并促进了植物免疫系统。植物细胞壁是坏死性病原体的主要碳源之一。通过植物细胞中吸收这些障碍并吸收了进一步的养分,病原体通过分泌切丁蛋白酶和其他细胞壁降解酶来吞噬其宿主。通过对细胞降解产物的感知(称为损伤相关的分子模式(DAMP))监测细胞壁的完整性,从而激活免疫反应[18]。潮湿是指在正常情况下通常不存在的分子,包括细胞壁成分,核酸片段,

BMP4 触发指定心脏中胚层谱系的调节电路
© 2023. 由 The Company of Biologists Ltd. 出版。这是一篇开放获取的文章,根据知识共享署名许可条款分发(http://creativecommons.org/licenses/by/4.0),允许在任何媒体中不受限制地使用、分发和复制,前提是原始作品得到适当的署名

事件触发者和意见领导者塑造了气候变化论述
了解现实世界中的事件和意见领导者如何塑造气候变化讨论对于改善沟通和政策制定以实现全球碳减少目标至关重要。这项研究分析了中国最大的社交媒体平台的微博(2012 - 2022)的530万个原始帖子,以研究气候变化话语。我们发现五个事件类型触发了48个讨论峰,包括在线活动,国际会议,极端天气,国内政策和国际新闻。帖子通常传达了积极的态度,尽管在阴霾污染和19009年大流行期间的情绪下降。网络分析揭示了具有不同策略的七个意见团体:官方媒体和机构强调政治意志,全球倡议和社会经济的影响,而大学和基层人物则专注于科学现实和个人行动。名人和非官方叙述通常强调地缘政治话题,尤其是中国关系。我们建议减少零散的回声室,并通过数字媒体平台促进个人联系,以增强公众意识。
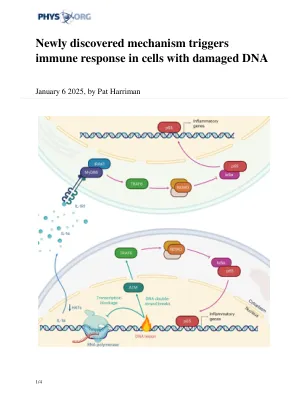
新发现的机制触发DNA受损细胞中的免疫反应
团队成员开发了一种先进的成像技术,以分析在细胞水平上如何调节NF-κB。研究人员能够精确地测量单细胞水平上细胞对受损DNA的反应,并观察到NF-κB激活的新途径。他们发现在特定类型的损伤后,细胞释放IL-1α蛋白。它不作用于细胞本身,而是传播到相邻细胞,在那里它触发了IRAK1蛋白,然后启动NF-κB炎症反应。

诱导物的父本染色体消除引发油菜双单倍体的诱导
合成的八倍体油菜籽 Y3380 在用作花粉供体为植物授粉时可诱导母本双单倍体。但双单倍体形成的潜在机制仍不清楚。我们推测双单倍体诱导发生在诱导系的染色体传递到母本卵细胞,并通过受精形成合子时。在合子有丝分裂过程中,父本染色体被特异性地消除。在消除过程中,部分父本基因可能通过同源交换渗入母本基因组。然后,合子单倍体基因组加倍(早期单倍体加倍,EH 现象),加倍的合子继续发育成完整的胚胎,最终形成双单倍体后代。为了验证假设,本研究以八倍体Y3380品系为标记,将4122-cp4-EPSPS外源基因回交,得到六倍体Y3380-cp4-EPSPS作为父本材料,对3个不同的母本材料进行授粉。在授粉后48 h观察诱导品系与母本杂交的受精过程,受精率分别达到97.92%和98.72%。授粉12 d后,用原位PCR检测胚中存在cp4-EPSPS,授粉后13 — 23 d,F 1 胚含有cp4-EPSPS基因的概率高达97.27%,而后逐渐下降,在23 — 33 d时为0%。同时免疫荧光观察了3~29天胚胎中cp4-EPSPS的表达情况。随着胚胎的发育,cp4-EPSPS标记基因不断丢失,伴随胚胎死亡,30天后在存活的胚胎中检测不到cp4-EPSPS的存在。同时对诱导后代的SNP检测证实了双单倍体的存在,进一步表明诱导过程是由于父本染色体特异性的丧失引起的。四倍体诱导后代表现出诱导系基因位点的筛选,有杂合性,也有纯合性。结果表明,在诱导过程中,诱导系染色体被消除。

神经影像学时间序列中的箭头 - 标识脑功能的因果触发器
图4箭头识别运动任务中的时空定位因果效应。(a)在运动任务范式中,因果效应(τ,顶部),活动(中间)和连通性(底部)的度量。范式由运动时期(左右手和脚,舌头)组成,被休息块隔开。(b)左半球大脑区域的因果效应的详细视图,显示了面板(a)(舌运动)突出显示的间隔中最强的AOT波动。正值表明该区域充当因果效应的下水道,而负值表明该区域是因果关系的来源。(c)面板(b)中四个大脑区域的可视化以及当受试者开始移动舌头时招募的假定因果途径。VIS24和PFC13之间的虚线表示,这两个区域之间的直接信息流不能仅从分析的四个区域中推断出来,并且可能涉及中间体。

神经祖细胞中的同时NBS1和p53失活触发高级神经胶质瘤
f i g u r e 1 p53失活挽救NBS1 NES-CRE有害脑表型。(a)通过p53失活在p21处拯救NBS1 NES-CRE脑缺陷。(b)与NBS1 NES-CRE EGL和大脑皮层相比,NBS1 NES-CRE,P53 / EGL和大脑皮层缺乏凋亡。比例尺=20μM。(c)与NBS1 NES-CRE的大脑相比,NBS1 NES-CRE EGL中的Tunel阳性细胞数量显着减少。nbs1 nes-cre(n = 3),nbs1 nes-cre,p53 /(n = 2),nbs1 ctrl(n = 4)。nbs1 nes-cre vs nbs1 ctrl(脑皮质**:p = 0.0018,egl ****:p <0.0001),nbs1 nes-cre,p53 / vs nbs1 nbs1 nbs1 nes-cre(大脑皮层NBS1 CTRL(脑皮质 *:P = 0,0181,EGL *:P = 0.0360)。(d)NBS1 NES-CRE和NBS1 NES-CRE,P53 / EGL表现出γ-H2AX灶。比例尺=20μM。(E)NBS1 NES-CRE和NBS1 NES-CRE,p53 / eGL和脑皮质中γ-H2AX +细胞的定量。n.s:没有显着差异。nbs1 nes-cre

使用模块化生态系统的自主发现在LHC触发器中进行自适应异常检测
异常检测(AD)代表了一种从根本上进行数据驱动发现的新工具。最初的努力集中在将强大的离线算法调整到这些高通量流系统中,但这种算法应如何适应不断发展的检测器条件的问题仍然是一个重大挑战。在这项工作中,我们引入了一个模块化生态系统,以制定和评估自主发现的策略,其中包含了不同的组件,包括:具有时间依赖性效果的数据集,复杂的触发菜单,实时控制机制和成本感知的优化标准。我们通过使用公共CMS数据集的AD触发器进行了基于强化学习的新基准来说明这一框架,旨在鼓励以社区为导向的发展发展新一代智能和适应性触发器。