XiaoMi-AI文件搜索系统
World File Search Systemviral

细胞抵御病毒感染的保护屏障 - Refubium
粘液是一种动态生物水凝胶,主要由糖蛋白粘蛋白组成,具有独特的生物物理特性,并形成保护细胞免受多种病毒侵害的屏障。在这里,这项工作开发了一种基于聚甘油硫酸盐的树枝状粘蛋白启发共聚物 (MICP-1),其中约 10% 的活性二硫化物重复单元作为交联位点。MICP-1 的低温电子显微镜 (Cryo-EM) 分析揭示了细长的单链纤维形态。MICP-1 对许多病毒表现出潜在的抑制活性,例如单纯疱疹病毒 1 (HSV-1) 和 SARS-CoV-2(包括 Delta 和 Omicron 等变体)。MICP-1 使用线性和支链聚乙二醇硫醇 (PEG-thiol) 作为交联剂,生产出具有与健康人痰液相似的粘弹性能和可调节微结构的水凝胶。使用单粒子跟踪微流变学、电子顺磁共振 (EPR) 和低温扫描电子显微镜 (Cryo-SEM) 来表征网络结构。合成的水凝胶表现出自修复特性,以及可通过还原调节的粘弹性能。使用 transwell 测定法来研究水凝胶对 HSV-1 病毒感染的保护特性。活细胞显微镜证实,由于网络形态和阴离子多价效应,这些水凝胶可以通过捕获病毒来保护底层细胞免受感染。总体而言,这种新型粘蛋白共聚物可生成数克级的粘液模拟水凝胶。这些水凝胶可用作富含二硫化物的气道粘液研究的模型,也可用作生物材料。

肝与先天免疫 - 病毒感染中的朋友或敌人
研讨会准备的出版物:Liang Y,Luo X,Schefczyk S,Muungani LT,Deng H,Wang B,Baba HA,Lu M,Lu M,Wedemeyer H,Schmidt HH,Broing r。乙型肝炎表面抗原表达会通过减少LAMP2而损害内质网应激相关的自噬通量。JHEPREP。2024JAN 28; 6(4):101012。 Schefczyk S,Luo X,Liang Y,Hasenberg M,Walkenfort B,Trippler M,Schuhenn J,Sutter K,Lu M,Wedemeyer H,Schmidt HH,Broing r。 TG1.4HBV-S-REC小鼠是一种杂交丙型肝炎病毒 - 转基因模型,发展为轻度的肝炎。 SCIREP。202312月20日; 13(1):22829。 luo X,Zhang R,Schefczyk S,Liang Y,Lin SS,Liu S,Baba HA,Lange CM,Wedemeyer H,Lu M,Bro.R. YAP的核转运驱动BMI相关的肝炎病毒BMI相关肝癌发生。 肝脏Int。 2023年9月; 43(9):2002-2016。 Luo X,Zhang R,Lu M,Liu S,Baba HA,Gerken G,Wedemeyer H,Broing R。 河马途径反调节丙型肝炎病毒感染中的先天免疫力。 前疫苗。 2021 5月25日; 12:684424。 Zhang Z,Trippler M,Real CI,Werner M,Luo X,Schefczyk S,Kemper T,Anastasiou OE,Ladiges Y,Treckmann J,Paul A,Baba HA,Allweiss L,Allweiss L,Dandri M,Dandri M,Dandri M,Gerken G,Gerken G,Gerken G,Gerken G,Wedemeyer H,Schlaak Jf,Schlaak Jf,laus jf,broim broive broim thiviv thiviv tiviv tiviv。受体2原发性肝细胞感染后的信号传导。 Hepatology 2020年9月; 72(3):829-844。 Broing R,Zhang X,Kottilil S,Trippler M,Jiang M,Lu M,Gerken G,Schlaak JF。 干扰素刺激的基因15是丙型肝炎病毒的前病毒因子,也是IFN反应的调节剂。 肠道。JHEPREP。2024JAN 28; 6(4):101012。Schefczyk S,Luo X,Liang Y,Hasenberg M,Walkenfort B,Trippler M,Schuhenn J,Sutter K,Lu M,Wedemeyer H,Schmidt HH,Broing r。TG1.4HBV-S-REC小鼠是一种杂交丙型肝炎病毒 - 转基因模型,发展为轻度的肝炎。SCIREP。202312月20日; 13(1):22829。 luo X,Zhang R,Schefczyk S,Liang Y,Lin SS,Liu S,Baba HA,Lange CM,Wedemeyer H,Lu M,Bro.R. YAP的核转运驱动BMI相关的肝炎病毒BMI相关肝癌发生。 肝脏Int。 2023年9月; 43(9):2002-2016。 Luo X,Zhang R,Lu M,Liu S,Baba HA,Gerken G,Wedemeyer H,Broing R。 河马途径反调节丙型肝炎病毒感染中的先天免疫力。 前疫苗。 2021 5月25日; 12:684424。 Zhang Z,Trippler M,Real CI,Werner M,Luo X,Schefczyk S,Kemper T,Anastasiou OE,Ladiges Y,Treckmann J,Paul A,Baba HA,Allweiss L,Allweiss L,Dandri M,Dandri M,Dandri M,Gerken G,Gerken G,Gerken G,Gerken G,Wedemeyer H,Schlaak Jf,Schlaak Jf,laus jf,broim broive broim thiviv thiviv tiviv tiviv。受体2原发性肝细胞感染后的信号传导。 Hepatology 2020年9月; 72(3):829-844。 Broing R,Zhang X,Kottilil S,Trippler M,Jiang M,Lu M,Gerken G,Schlaak JF。 干扰素刺激的基因15是丙型肝炎病毒的前病毒因子,也是IFN反应的调节剂。 肠道。SCIREP。202312月20日; 13(1):22829。luo X,Zhang R,Schefczyk S,Liang Y,Lin SS,Liu S,Baba HA,Lange CM,Wedemeyer H,Lu M,Bro.R. YAP的核转运驱动BMI相关的肝炎病毒BMI相关肝癌发生。肝脏Int。 2023年9月; 43(9):2002-2016。 Luo X,Zhang R,Lu M,Liu S,Baba HA,Gerken G,Wedemeyer H,Broing R。 河马途径反调节丙型肝炎病毒感染中的先天免疫力。 前疫苗。 2021 5月25日; 12:684424。 Zhang Z,Trippler M,Real CI,Werner M,Luo X,Schefczyk S,Kemper T,Anastasiou OE,Ladiges Y,Treckmann J,Paul A,Baba HA,Allweiss L,Allweiss L,Dandri M,Dandri M,Dandri M,Gerken G,Gerken G,Gerken G,Gerken G,Wedemeyer H,Schlaak Jf,Schlaak Jf,laus jf,broim broive broim thiviv thiviv tiviv tiviv。受体2原发性肝细胞感染后的信号传导。 Hepatology 2020年9月; 72(3):829-844。 Broing R,Zhang X,Kottilil S,Trippler M,Jiang M,Lu M,Gerken G,Schlaak JF。 干扰素刺激的基因15是丙型肝炎病毒的前病毒因子,也是IFN反应的调节剂。 肠道。肝脏Int。2023年9月; 43(9):2002-2016。Luo X,Zhang R,Lu M,Liu S,Baba HA,Gerken G,Wedemeyer H,Broing R。河马途径反调节丙型肝炎病毒感染中的先天免疫力。前疫苗。2021 5月25日; 12:684424。Zhang Z,Trippler M,Real CI,Werner M,Luo X,Schefczyk S,Kemper T,Anastasiou OE,Ladiges Y,Treckmann J,Paul A,Baba HA,Allweiss L,Allweiss L,Dandri M,Dandri M,Dandri M,Gerken G,Gerken G,Gerken G,Gerken G,Wedemeyer H,Schlaak Jf,Schlaak Jf,laus jf,broim broive broim thiviv thiviv tiviv tiviv。受体2原发性肝细胞感染后的信号传导。Hepatology 2020年9月; 72(3):829-844。Broing R,Zhang X,Kottilil S,Trippler M,Jiang M,Lu M,Gerken G,Schlaak JF。干扰素刺激的基因15是丙型肝炎病毒的前病毒因子,也是IFN反应的调节剂。肠道。2010年8月; 59(8):1111-9。

组蛋白脱乙酰基酶6在病毒感染中的功能,先天...
在组蛋白二乙酰酶家族中,组蛋白脱乙酰基酶6(HDAC6)脱颖而出。细胞质IIB类组蛋白脱乙酰基酶(HDAC)家族对于许多细胞功能至关重要。它在先天抗病毒免疫中起着至关重要且有争议的调节作用。本综述总结了我们对HDAC6控制DNA和RNA病毒感染的三种机制的理解的当前状态:细胞骨架调节,宿主先天免疫反应以及宿主或病毒蛋白的自噬降解。此外,我们总结了HDAC6抑制剂如何用于治疗多种疾病,以及其上游信号如何在抗病毒机制中起作用。,这篇综述的发现重点介绍了HDAC6在抗病毒免疫,先天免疫反应和某些疾病方面的新治疗靶标的重要性,所有这些疾病都为针对免疫反应的药物开发提供了有希望的新途径。

DNA结合重新分布激活域的集合和先锋因子Sox2各种病毒cas基因拮抗CRISPR免疫
(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。该预印本版的版权持有人于2023年6月24日发布。 https://doi.org/10.1101/2023.06.24.545427 doi:biorxiv preprint


欧洲的肝癌不平等和病毒肝炎的作用
据估计,欧盟和欧洲经济区30个国家(EU27/EEA)的人口中约有1.0%的人口患有慢性丙型肝炎感染,患病率估计从爱尔兰的0.1%到罗马尼亚的4.5%不等(4,5)。欧盟国家慢性丙型肝炎感染的平均患病率估计为0.5%,范围从荷兰和斯洛文尼亚的0.1%到罗马尼亚的2.3%(6)。然而,世界卫生组织估计,全球87%的丙型肝炎和64%的丙型肝炎感染未被诊断(7)。建模的数据表明,在EU27/EEA国家/地区,未诊断的丙型肝炎病例的比例从丹麦的0%到匈牙利的95%,以及未诊断的乙型肝炎病例的比例范围从西班牙和挪威的0%到葡萄牙的97%到葡萄牙的97%,大多数国家都有最高的疾病攻击。

记录或操纵昆虫神经元的遗传和病毒方法
用于记录和操纵体内神经元的遗传编码工具的发展极大地提高了我们对神经元活动如何影响行为的理解。最近的进展使得这些工具可用于通常被认为不易被遗传处理的物种。这一进展正在彻底改变神经科学,特别是昆虫神经行为学。这里我们介绍了最新的创新及其在系统发育多样化的昆虫物种中的一些应用。我们讨论了这些方法对基础研究和转化研究的重要性和意义。我们重点介绍用于钙成像、光遗传学和突触沉默的遗传编码和病毒编码工具。最后,我们讨论了用于昆虫行为神经行为学研究的普遍适用、模块化和用户友好的遗传工具包的未来潜在发展。

靶向病毒CCCDNA治愈慢性乙型肝炎
审查的抽象目的是由乙型肝炎病毒(HBV)引起的慢性肝炎B(CHB)是全球晚期肝病和肝细胞癌(HCC)的主要原因。HBV复制的特征在于合成共价闭合(CCC)DNA,而抗病毒核(T)IDE类似物(NUC)的靶向是护理标准标准的关键方式。尽管HBV复制成功地抑制了经过治疗的患者,但他们仍有患HCC的风险。虽然功能治疗(以HBSAG损失为特征)是新型抗病毒药疗法的第一个目标,但消除CCCDNA的治疗疗法仍然是最终目标。本评论总结了新型治疗策略的发现和发展的最新进展及其对CCCDNA生物学的影响。最近十年的最新发现,在理解CCCDNA生物学方面取得了重大进展,包括发现宿主依赖性因素,CCCDNA转录的表观遗传调节和免疫介导的降解。目前以直接或间接方式针对CCCDNA的几种方法正在发现,临床前或早期临床发育的阶段。示例包括基因组编辑方法,针对宿主依赖性因素或表演基因调控的策略,核素蛋白质调节剂和免疫介导的降解。总结直接定位的CCCDNA策略仍处于开发的临床前阶段,但带帽子装配调节器和基于免疫的方法已经达到了临床阶段。组合疗法提供了更多机会来克服当前方法的局限性。临床试验正在进行评估其对患者的功效和安全性,包括对病毒CCCDNA的影响。

ASHP政策职位1982-2024
美国卫生系统药剂师协会公司表示,有关Janssen(Johnson and Johnson)Covid-19疫苗的此信息是配备了合理的护理标准,并符合该领域的专业标准。读者被告知,Janssen(Johnson and Johnson)Covid-19疫苗不是SARS-COV-2引起的冠状病毒病2019(COVID-19)的批准疫苗,而是正在调查,目前正在调查,目前已在FDA紧急使用授权(EUA)下可用来防止某些成年成年人in Covid-19。美国卫生系统药剂师协会公司不对明示或暗示的任何陈述或保证,包括但不限于对特定目的的适销性和适合性的任何暗示保证,在信息方面,有关信息,并具体否认所有此类保证。有关Janssen(Johnson and Johnson)Covid-19疫苗的信息的读者,建议ASHP对信息的持续货币,任何错误或遗漏和/或因使用此信息而引起的任何后果概不负责。读者被告知,有关药物治疗的决定是需要复杂的医疗决定,需要适当的医疗保健专业人员的独立,明智的决定,并且仅出于信息目的提供了此信息中包含的信息。美国卫生系统药剂师协会不认可或建议使用任何药物。有关Janssen(Johnson and Johnson)Covid-19疫苗的此信息不应被视为个体的患者建议。由于药物信息的性质变化,建议您咨询您的医师或药剂师有关任何药物的特定临床使用。
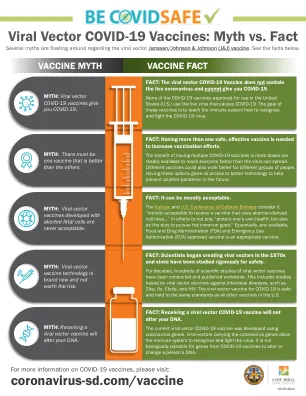
病毒载体covid-19疫苗:神话与事实
梵蒂冈和美国天主教主教会议认为,“在道德上接受一种使用堕胎衍生的细胞系的疫苗……在道德上可以接受……”,不仅要“保护自己的健康,而且还有责任追求共同利益”。本质上,任何可用的食品药品监督管理局(FDA)和紧急使用授权(EUA)批准的疫苗都是适当的疫苗。