XiaoMi-AI文件搜索系统
World File Search System不对称的

不对称反奈定向光毒素催化
10。L. J. Rono,H。G. Yayla,D。Y. Wang,M。F. Armstrong,R。R. R. Knowles,proton耦合电子传递启用了对映射光介毒催化:开发不对称的AZA AZA-PINACOL环化。j。am。化学。Soc。135,17735–17738(2013)。

与伊拉克政府规模有关的经济增长
(1995)。换句话说,政府支出的增加是有益的,直至一定的阈值,但是超过该水平,对增长的影响是负面的。从经验测试条形曲线的研究包括:Karra(1997),Chen和Lee(2005),Chiou-Wei等。(2010),Gunalp和Dincer(2010)以及Altunc andAydın(2013)。 为了建立最佳的政府支出水平,应采用丰富的国家小组。 最重要的是,存在一个普遍的内生性问题,在文献中尚未解决,因为一段时间内较高的增长可能会促使政府通过税收较高的回报渠道促进更大的支出(4)。 表明,即使我们将样本分为发达国家和发展中国家,这一影响仍然有效,对这两组有不对称的影响(5)。(2010),Gunalp和Dincer(2010)以及Altunc andAydın(2013)。为了建立最佳的政府支出水平,应采用丰富的国家小组。最重要的是,存在一个普遍的内生性问题,在文献中尚未解决,因为一段时间内较高的增长可能会促使政府通过税收较高的回报渠道促进更大的支出(4)。表明,即使我们将样本分为发达国家和发展中国家,这一影响仍然有效,对这两组有不对称的影响(5)。

使用脑MRI MRI
图1在疾病的早期和晚期阶段,具有不同SCJD亚型患者的代表性扩散加权图像。(第一行)具有MM1亚型的患者的早期DWI,该患者在左顶叶皮层(包括前后节)和前额叶皮质丝带中表现出高强度。不同的MM1患者的晚期DWI显示出左脑半球的大多数皮质的不对称参与,与尾状头部,左扣带回和左岛群体结合。(第二行)患有MM2C亚型的患者的早期DWI,在左侧顶叶皮层中出现超强度,包括前神经。不同MV2C患者的晚期DWI显示皮质色带不对称受累;纹状体和丘脑被幸免。(第三行)患有VV1亚型的患者的早期DWI,在左顶皮层中表现高强度,左扣带回和绝缘。不同VV1患者的晚期DWI表现出大脑皮层和右纹状体的广泛不对称受累。thalami和大多数左脑皮质都幸免了。(第四行)MV2K亚型患者的早期DWI,他在纹状体的纹状体和微妙的双侧超强度中表现出了不对称的高强度。不同的MV2K患者的晚期DWI表现出纹状体和整个丘脑中不对称的高强度,以及左额叶皮层和岛菌的轻度参与。顶层皮质幸免。(最后一行)患有VV2亚型患者的早期DWI,在尾状的头部和壳质的前部出现了不对称的DWI高强度,以及丘脑中非常微妙的高强度。具有VV2的不同患者的晚期DWI显示了纹状体和丘脑的广泛超强度,以及左回扣和前额叶皮层。scjd,零星的克鲁特兹菲尔特 - 贾科布疾病; DWI,扩散加权成像。

绘制大脑不对称图
绘制大脑不对称图谱 Arthur W. Toga 和 Paul M. Thompson 神经成像实验室 加州大学洛杉矶分校医学院神经病学系 摘要 动物和人类都存在大脑结构、功能和行为上的不对称现象。这种偏侧性被认为源于进化、遗传、发育、经验和病理因素。本文回顾了描述大脑不对称的各种文献,主要关注那些描述大脑半球解剖学差异的观察结果。 简介 大多数生物系统都表现出一定程度的不对称 1 。从人类到低等动物,正常的变异和特化都会产生功能和结构的不对称。甚至面部和四肢的外部特征也能证明这种不对称 2 。在人类和许多其他哺乳动物中,两个大脑半球在解剖学和功能上有所不同。虽然粗略检查人脑的宏观特征无法发现明显的左右差异,但仔细检查其结构就会发现各种不对称特征。这种侧化特化被认为源于进化、发育、遗传、经验和病理因素。例如,左半球语言皮层的进化扩张可能导致布罗卡言语区、颞平面(颞叶后部的听觉处理结构)和其他对言语产生、感知和运动优势至关重要的结构出现明显的体积不对称。大脑功能布局、细胞结构和神经化学的不对称也与不对称行为特征有关,例如惯用手、听觉感知、运动偏好和感觉敏锐度。在这里,我们回顾了各种方法及其对大脑结构和功能不对称的观察结果,特别关注解剖学差异。特别是大脑映射方法可以检测和可视化整个人群的不对称模式,包括疾病、年龄和发育过程中的细微变化。这些工具和其他工具在评估调节大脑认知特化的因素方面显示出巨大的潜力,包括大脑不对称的个体发育、系统发育和遗传决定因素。语言和惯用手语言。左半球的语言特化是大脑不对称的最早观察结果之一。19 世纪的 Broca 3 和 Wernicke 4 报告称,语言会因左半球的肿瘤或中风而受到更严重的损害。语言产生和句法处理的某些方面 5,6 随后主要定位到左半球前部的区域,包括下额回的三角部和岛叶部(布罗卡区;见图 1)。另一方面,语言理解,例如理解口语 7 ,主要局限于后颞顶叶区域,包括韦尼克区(布罗德曼区 39、40、后 21 和 22 以及 37 的一部分)。许多行为任务进一步阐明了语言回路,包括语法处理、语义知识和句法测试 8、5-6、9 。

补充信息压力引起的...
图 S1 显示了使用 Amam 和 Fmmm 空间群在 295 K 下测量的 La 3 Ni 2 O 7-δ 多晶样品的 NPD 图案的 Rietveld 细化结果比较。尽管对数据的细化似乎相似,但只有 Amam 对称性才允许反射。2 θ 区域从 40º 到 50º 的 NPD 数据也显示出不对称的 Warren 类峰形,这通常与短程有序有关。
神圣的心学院(自主)
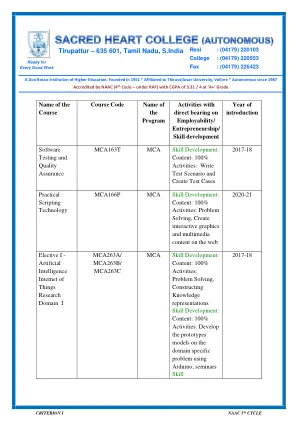
MCA466A/MCA466B MCA技能开发:内容:100%活动:应用传递数据所需的技术和软件包,并可视化它们,将各种统计/学习技术应用于给定问题技能开发的数据分析中的各种统计/学习技术:内容:100%活动:100%活动:Quiz,Quiz,在密码,对称性和不对称的cipsography,Symmetric和symmetric cip cip cip cip cip cip cip cip

映射新兴技术及其供应链
新兴技术对政策制定者,私人公司和研究人员的兴趣浓厚。这种兴趣源于共同的看法,即在生物技术,人工智能/机器学习(AI/ML),量子计算和先进的古典计算等关键领域的进步将在未来几十年中赋予不对称的经济和国家安全优势。但是,正在进行的供应链中断暴露了某些技术全球价值链的全球依赖和脆弱性。


生物膜结构和成分-II的流动性和不对称性:目标:本讲座的主要目标是:简要讨论不规则
细胞的边界是由生物膜形成的,即定义细胞内部和外部的屏障。这些障碍可以防止细胞内部产生的分子泄漏出来,并从扩散中散开分子;然而,它们还包含允许细胞采用特定分子并去除不需要的传输系统。此类运输系统授予膜选择性渗透性的重要特性。膜是动态结构,其中蛋白质漂浮在脂质的海中。膜的脂质成分形成了通透性的屏障,蛋白质成分充当泵和通道的传输系统,可将选定的分子进入和流出细胞。生物膜形成不对称结构,并且像具有流动性一样是流体,即具有各种细胞分子的易位酶。生物膜的不对称性可以部分归因于膜内蛋白质的不规则分布。生物膜的脂质双层由外部小叶和内部小叶组成,它们分布在两个表面之间,以在外表面和内表面之间形成不对称性。这个不对称的组织对于细胞功能(例如细胞信号传导)很重要。生物膜的不对称性反映了膜的两个传单的不同功能。如磷脂双层的流体膜模型所示,膜的外部和内部小叶在其组成中是不对称的。膜流动性是指

9/14/2020 1“差分目标多路复用”脊髓...
相关图说明了基因表达,SCS电流(MA)和行为评分(BSPB)的百分比之间的关系。a:sham,b:no-scs(sni),C:双相对称SCS,D:单相阴性SCS,E:单相阳极SCS,F:非对称性双相SCS 1:2,G:不对称的双偶联1:0.5。蓝点代表正相关,红点代表负相关。点的大小和黑暗与Pearson相关系数的值成正比