XiaoMi-AI文件搜索系统
World File Search System中端


拓扑层结构中的光学双稳态及其在光神经网络中的应用*
材料Sio 2。在拓扑模式下,电场高度局部位于分层结构的反转中心(也称为界面),并成倍地衰减到批量上。因此,当从战略上引入非线性介电常数时,出现了非线性现象,例如Biscable状态。有限元数值模拟表明,当层周期为5时,最佳双态状态出现,阈值左右左右。受益于拓扑特征,当将随机扰动引入层厚度和折射率时,这种双重状态仍然存在。最后,我们将双态状态应用于光子神经网络。双态函数在各种学习任务中显示出类似于经典激活函数relu和Sigmoid的预测精度。这些结果提供了一种新的方法,可以将拓扑分层结构从拓扑分层结构中插入光子神经网络中。

终端管制技术交流会议
壹、目的 ............................................................................................................................... 3

基于近端策略优化算法及视觉感知的机械臂导纳控制研究
在现代操纵器交互任务中,由于环境的复杂性和不确定性,准确的对象表面建模通常很难实现。因此,改善操纵器与环境之间相互作用的适应性和稳定性已成为相互作用任务的重点之一。针对操纵器的互动任务,本文旨在在视觉指导下实现良好的力量控制。因此,基于Mujoco(带有触点的多关节动力学)物理引擎,我们为操纵器构建了交互式仿真环境,并创新地集成了基于位置的视觉伺服控制和录取控制。通过深度强化学习(DRL)中的近端策略优化(PPO)算法,有效地集成了视觉信息和力量信息,并提出了结合视觉感知的接收性控制策略。通过比较实验,将允许控制与视觉感知相结合,并将力控制的整体性能提高了68.75%。与经典的入学控制相比,峰值控制精度提高了15%。 实验结果表明,在平坦和不规则的凹面环境中,允许控制与视觉感知结合表现良好:它不仅可以准确地执行视觉构成的力控制任务,而且还可以在各种接触表面上维持施工力,并迅速适应环境变化。与经典的入学控制相比,峰值控制精度提高了15%。实验结果表明,在平坦和不规则的凹面环境中,允许控制与视觉感知结合表现良好:它不仅可以准确地执行视觉构成的力控制任务,而且还可以在各种接触表面上维持施工力,并迅速适应环境变化。在精确组装,医疗援助和服务操纵器的领域中,它可以提高操纵器在复杂和不确定的环境中的适应能力和稳定性,从而促进智能操纵器的自主操作的发展。

人工智能在油气钻井工程中的应用
进入石油和天然气钻井工程,改善了钻井操作的智能水平。根据国内外石油和天然气钻探工程的当前研究状况,本文讨论了人工智能在石油和天然气钻探工程中的关键技术应用。智能钻井和完成技术结合了大数据,人工智能算法和软件平台,以优化关键技术,例如井眼轨迹,定向钻孔和钻孔速度,以提高操作安全性和效率。其次,智能钻井设备的研发和应用在国际上是相对成熟的。诸如智能钻机,钻头和旋转转向系统之类的设备已经达到了高度的自动化,从而提高了运行效率并降低了人工成本。最后,钻井和完成软件系统通过引入机器学习和云计算等技术来集成和分析大量数据,从而优化了钻孔设计和操作。尽管中国在智能钻探软件和设备领域开始后期开始,但它取得了一些进展,主要是在监视优化和设计方面。将来,随着核心技术的突破,人工智能将为石油和天然气资源的发展带来一场技术革命。中国需要继续加强基础研究,结合行业的实际需求,并促进独立的技术创新和应用促进,以提高智力的整体水平,并通过国际高级技术来缩小差距。

2014 CSR - 中华航空
华航成立于1959年,即将迈入第56个年头。「留住满意的顾客与快乐的员工,为股东与社会创造最大价值」一直是华航自成立以来的经营理念。华航致力成为最值得信赖的世界级航空公司,以最佳的飞行品质,让每一位旅客都感到满意。截至2015年6月30日,华航客货机队规模已扩展至86架,航线网络覆盖全球29个国家115个航点。去年,在大家的共同努力下,华航及其关联企业交出亮眼的成绩单。2015年,受国际油价平稳、日圆贬值、大陆旅客经台过境限制即将解除等因素影响,旅客量持续增长,货运市场也逐渐回暖。感谢每一位华航员工、旅客、股东及伙伴的支持与鼓励。

糖尿病与卒中后抑郁的研究进展
中风后抑郁症是中风的常见并发症,严重影响了患者的生活和生活质量。大约三分之一的中风患者被诊断出患有糖尿病,糖尿病会加剧卒中后抑郁症的风险。在病理生理学方面,两者之间存在相互加强的关系。本文将探讨糖尿病与中风后抑郁症之间的病理生理联系,并从药物疗法的角度进行分析,旨在为也患有糖尿病的卒中后抑郁症患者提供预防和治疗参考。

强于大市AI 端侧深度报告之AI 手机
智能手机是最适合承载端侧 AI 的载体, AI 手机可提供差异化的用户价 值与品牌价值。智能手机具有保有量大、使用便携、使用场景多、使用 时长久、应用生态系统强大等优势,可创造众多的 AI 使用场景,并加速 第三方 AI 应用成熟,我们认为智能手机将是生成式 AI 最佳的应用载体 之一。 AI 手机的定义具有三个典型特征:①能够在手机端侧运行大模型; ② SoC 中包含 NPU 算力;③达到一定参数要求的性能指标。 AI 手机可提 供差异化的用户价值与品牌价值。对用户而言, AI 手机将是自在交互、 智能随心、专属陪伴、安全可信的个人化助理,使用体验较目前阶段智 能手机大幅提升。对于手机厂商而言,可提供品牌形象与用户粘性。

讯飞AI 考试系统学生手机端使用说明
1.下载 ............................................................................................................................................... 3
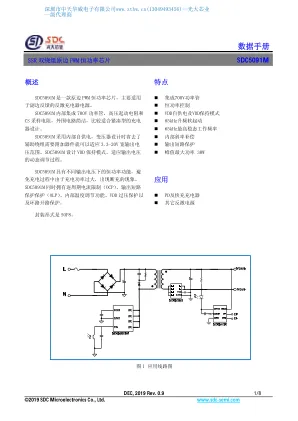
数据手册 - 中天华威
2019年12月Rev.0.9 1/8©2019 SDC Microelectronics Co.,Ltd。www.sdc-semi.com0.9 1/8©2019 SDC Microelectronics Co.,Ltd。www.sdc-semi.com