XiaoMi-AI文件搜索系统
World File Search System之树

片上集成人工微结构光场调控(特邀) - 陈树琪
[20] Liu W W,Chen S Q,Li Z C等。使用单层跨表面[J]在Terahertz区域中在Terahertz区域中传输模式下的极化转换实现。光学信,2015,40(13):3185-3188。

“AI+R”树:实例优化的 R 树
摘要 — 新兴的实例优化系统类别已显示出通过专门针对特定数据和查询工作负载实现高性能的潜力。特别是,机器学习 (ML) 技术已成功应用于构建各种实例优化组件(例如,学习索引)。本文研究了利用 ML 技术来增强空间索引(特别是 R 树)的性能,以适应给定的数据和查询工作负载。由于 R 树索引节点覆盖的区域在空间中重叠,因此在搜索空间中的特定点时,可能会探索从根到叶的多条路径。在最坏的情况下,可以搜索整个 R 树。在本文中,我们定义并使用重叠率来量化范围查询所需的无关叶节点访问程度。目标是提高传统 R 树对高重叠范围查询的查询性能,因为它们往往会产生较长的运行时间。我们引入了一种新的 AI 树,将 R 树的搜索操作转换为多标签分类任务,以排除无关的叶节点访问。然后,我们将传统的 R 树扩展到 AI 树,形成混合的“AI+R”树。“AI+R”树可以使用学习模型自动区分高重叠查询和低重叠查询。因此,“AI+R”树使用 AI 树处理高重叠查询,使用 R 树处理低重叠查询。在真实数据集上的实验表明,“AI+R”树可以将查询性能提高到传统 R 树的 500% 以上。
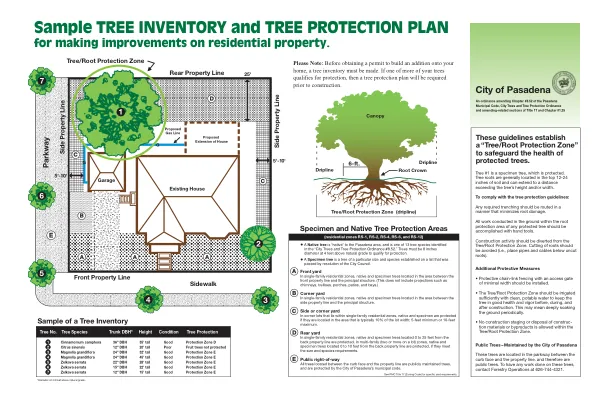
样本树清单和树保护计划
在单户住宅区的前院,本机和标本树位于前财产线和主要结构之间的区域。(这不包括诸如烟囱,格子,门廊,露台和海湾等预测。)在单户住宅区的角院,位于侧属性线和主要结构之间的区域的本地和标本树。

互动树生命树Itol v6
摘要生命之树(https://itol.embl.de)是用于管理,显示,注释和操纵系统发育和其他树木的在线工具。它是可以自由的,可以向E viry开放。Itol v ersion 6引入了现代化且完全重写的用户界面以及许多新功能。已经引入了一种新的数据集类型(彩色 /标记的范围),大大升级了先前的简单彩色范围注释函数的功能。对几个现有数据集T ypes实现了其他注释选项。DAT ASET模板文件现在通过子字符串匹配(包括完整的正则表达支持)来支持对多个树节点的简单分配。节点MET ADAT ADAT已大大扩展了处理,没有V el distai y和e Xporting选项,并且不能进行交互性编辑或通过注释文件进行更新。可以使用多个同时的字体样式显示树标签,并具有精确的定位,大小和单个标签零件的大小。实施了各种散装标签编辑功能,简化了所有树节点标签的大规模更改。ITOL的自动税收分配功能现在还基于基因组税元数据库(GTDB)支持树,此外NCBI税收税也是如此。可选的用户帐户页面的功能已扩展,简化了项目和树木的管理,导航和共享。ITOL目前从> 130 0 0 0单个用户帐户中处理超过一百万棵树。

科技树
6ROADS Adaptronica 空中客车波兰 Antmicro Astri Polska Astronika Blue Dot Solutions 天文中心。 MK PAN 空间研究中心 PAN Cervi Robotics CloudFerro Creotech Instruments Eversis 技术合作伙伴基金会技术合作伙伴 Geosystems GMV Innovating Solutions WB Group Hertz Systems Ltd ICEYE Polska InPhoTech 国家电信研究所 波兰科学院基础技术问题研究所 ITTI Jakusz Space Tech Kapitech Komes KP Labs Microamp Solutions N7 Space Nobo Solutions Opegieka PCO PIAP Space


树分解
4在图中的树中分类27 4.1图形定理的较弱版本。。。。。。。。。。。。。。。27 4.2网格和棕褐色。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。28 4.3不包括平面图。。。。。。。。。。。。。。。。。。。。。。。。。。。30 4.3.1分离和k -meshes。。。。。。。。。。。。。。。。。。。。。。31 4.3.2找到R -Grid未成年人。。。。。。。。。。。。。。。。。。。。。。。32 4.4有界树宽度的良好排序图。。。。。。。。。。。。。36 4.4.1对称的下函数和分支宽度。。。。。。。。37 4.4.2有界分支宽度的良好排序图。。。。。。。40 4.4.3将平面图排除为未成年人的含义。。。。。。。。42 4.5 kuratowski定理的概括。。。。。。。。。。。。。。。。。。43

科技树
6ROADS Adaptronica 空中客车波兰 Antmicro Astri Polska Astronika Blue Dot Solutions 天文中心。 MK PAN 空间研究中心 PAN Cervi Robotics CloudFerro Creotech Instruments Eversis 技术合作伙伴基金会技术合作伙伴 Geosystems GMV Innovating Solutions WB Group Hertz Systems Ltd ICEYE Polska InPhoTech 国家电信研究所 波兰科学院基础技术问题研究所 ITTI Jakusz Space Tech Kapitech Komes KP Labs Microamp Solutions N7 Space Nobo Solutions Opegieka PCO PIAP Space

量子默克尔树
提交信息是密码学的核心任务,其中一方(通常称为证明者)存储一段信息(例如,一个比特串)并承诺不更改它。另一方(通常称为验证者)可以访问此信息,后者可以稍后了解该信息并验证它没有被篡改。Merkle 树 [1] 是一种众所周知的简洁构造,其中验证者可以通过从诚实的证明者那里收到一个简短的证明来了解信息的任何部分。尽管 Merkle 树在古典密码学中具有重要意义,但却没有与 Merkle 树相关的量子类似物。直接使用量子随机预言模型(QROM)[2] 进行概括似乎并不安全。在这项工作中,我们提出了量子 Merkle 树。它基于我们所说的量子 Haar 随机预言模型(QHROM)。在 QHROM 中,证明者和验证者都可以访问 Haar 随机量子预言机 G 及其逆。利用量子 Merkle 树,我们为 Gap-k-Local-Hamiltonian 问题提出了一个简洁的量子论证。假设量子 PCP 猜想是正确的,这个简洁的论证可以扩展到所有 QMA 。这项工作提出了许多有趣的开放研究问题。
