XiaoMi-AI文件搜索系统
World File Search System传代

再生医学产品和...
a) 化学和药物信息 b) 细胞采集:供体筛选、同种异体和自体的测试和其他要求、组织分型{组织相容性抗原、组织分型过程、验收标准}、采集过程 c) 细胞/载体/靶基因序列的储存程序和方法:细胞/载体/靶基因序列的起源、来源、历史、细胞培养程序、传代控制、IPQC、表征主细胞/载体构建体/宿主细胞储存控制、表型/基因型表征等 3) 制造过程及其控制:a) 制造过程中所用材料的规格数据和经过验证的测试方法 b) 制造过程 c) 配方数据 d) 规格和质量控制 4) 表征:a. 细胞/GTP 身份 b. 细胞/GTP 纯度 c. 载体详细信息

hiPSC 中的敲除/SNP 敲入方案
基因工程将细胞置于选择压力之下,需要几轮细胞倍增才能获得编辑后的克隆。因此,为避免基因组不稳定性积累,我们建议使用解冻后 2-3 次传代的细胞,尽可能接近质量测试过的细胞库。我们还建议在缺氧条件下(37 C/5% CO 2 /5% O 2 )维护 hiPSC 并进行基因编辑实验,因为在缺氧条件下培养 hiPSC 有几个优点,包括增强多能性、增加增殖、减少氧化应激、提高重编程效率、更好的分化潜力和低遗传不稳定性频率。2、3 这些好处可以提高 hiPSC 的质量和功能,这对于再生医学和疾病建模中的下游应用至关重要。Vallone 等人描述了描述板涂层、细胞维护以及酶促和非酶促解离的一般方案。4

建立永生海洋鱼细胞系作为体外工具,促进环境监测、生物技术和生物多样性的研究
永生化细胞系对研究人员来说非常宝贵,因为它们可以无限增殖,从而可以培养多代。与寿命有限的原代细胞不同,永生化细胞系(也称为连续细胞系)可以避免伦理问题、提取困难、传代能力有限以及由于细胞来源不同而导致的结果不一致等挑战。实验室条件下的大多数细胞都面临海弗利克极限,即端粒随着每次分裂而缩短,导致衰老。永生化细胞系克服了这些限制,可以进行稳定的长期研究,同时无需重复分离和培养细胞,从而节省了研究的时间和资源。连续细胞系可以在体外无限增殖,为广泛的研究目的提供可持续和可重复的系统。这些细胞系为在细胞和分子水平上研究生物体提供了一个独特的平台,提供了全生物模型并不总是能够提供的见解。它复制了宿主的细胞和遗传同质性,同时最大限度地减少了体内系统固有的变异性。因此,人们越来越关注开发新的细胞系以支持更广泛的生物学研究。

利用 CRISPR/Cas9 在小鼠多能干细胞中进行快速、无整合的基因组编辑方法
CRISPR/Cas9 系统前所未有地革新了基因组编辑技术,该技术已成功应用于几乎所有生物科学分支。尽管在基因操作方面取得了很大成功,但大多数方法仍然费力且需要整合,并且需要长时间来扩增突变细胞库/克隆,而表现出功能性敲除效率的细胞较少。为了克服这些障碍,我们在此描述了一种高效、廉价、无整合且快速的一步式方案,用于小鼠多能干细胞 (PSC) 中的 CRISPR/Cas9 辅助基因敲除。我们的方案简化了基于脂质体的转染系统和筛选策略,使其能够更有效地处理少量 PSC(~2.0 × 10 4 个细胞),并最大限度地减少慢病毒包装、转导和单克隆传代等繁琐的步骤。在我们的方法中,约 90%(CI = 95%,79.5230% – 100%)的 PSC 菌落具有蛋白质表达方面的功能性敲除。因此,目前的方案在技术上可行、省时且高效,可用于多能干细胞中的基因组编辑。

13 干燥山羊疫苗于……被引入埃及。
1945 年,一种干燥山羊疫苗被引入埃及,作为对抗牛瘟再次入侵的大规模免疫方法,六个月后疫情被根除。山羊适应疫苗廉价有效,诱导的免疫力持久。然而,它们仍然存在低温保存的问题,即使通过干燥和真空储存提高了保存质量。在日本和韩国,兔子被用来适应病毒,用于对极易感染的牛品种进行血清同步疫苗接种(Nakamura 等人,1943 年)。经过多次传代,它仍然偶尔导致死亡,而在印度和蒙古牛中只发生轻微反应。1941 年,传代病毒在蒙古安全有效地使用,无需血清支持(Isogai,1944 年)。随后,兔化疫苗在非洲和亚洲广泛使用。 20 世纪 40 年代初,该疫苗在华北地区广泛使用,1945 年联合国善后救济总署在中国畜牧业研究局进一步研究,研制出一种疫苗(中村三号),该疫苗在牛和水牛身上只引起轻微反应。此后,随着和平的恢复,粮农组织将这种疫苗传播到埃及、泰国、印度、肯尼亚、巴基斯坦和埃塞俄比亚等许多国家(Hambidge 1955)。大约从 1950 年开始,哈尔滨兽医研究所的中国工作人员开始研制一种更令人满意的减毒活疫苗,因为中村三号疫苗难以按需要量生产。在兔、山羊和绵羊身上进行了数百次传代后,最终从淋巴结和脾脏中生产出一种疫苗,该疫苗对所有物种和品种都安全有效,甚至对牦牛和朝鲜牛也是如此。在牦牛身上,疫苗免疫持续时间经测试超过五年。这种疫苗被用于中国最后的根除行动(Roeder 等人,2006 年)。细胞培养技术的出现使工作人员能够将现有的减毒实验室牛瘟菌株改造成这种新基质,但直到 Walter Plowright 在牛肾细胞中培养出致命的 Kabete O 病毒 70 代后才取得突破;这种组织培养牛瘟疫苗 (TCRV) 既不产生病变也不产生发烧(Plowright,1962 年),并且对所有品种、年龄和性别的牛都是安全且具有免疫原性的。在日本,兔化/禽化 Nakamura III 病毒在 Vero 细胞中生长,以生产适用于日本牛的战略储备(Sonoda,1983 年)。同样,中国哈尔滨研究所目前正在羔羊肾细胞培养中生产中国兔化/山羊化/绵羊化疫苗的战略储备。 1961 年在喀布尔分离出一种致命的牛瘟病毒株,该病毒株在 37 次牛传代中得以保存,随后在哈萨克斯坦农业科学研究所的原代小牛肾细胞中进行了 70 次减毒,并于 1978 年作为疫苗推出。该疫苗在苏联与邻国之间的边境免疫带中常规使用,并在必要时用于抵御牛瘟的传入。这种疫苗被称为 K37/70,在评估期间经过了广泛的测试,随后被广泛使用,被认为对牛和牦牛是安全的。不幸的是,如前所述,在最近接种过这种疫苗的地区发生了临床牛瘟疫情。弗拉基米尔全俄动物健康研究所的科学家对 F 基因核苷酸序列(碱基 840 至 1161)进行了比较,结果表明 K37/70 病毒和喀布尔病毒只有一个碱基不同。此外,疫苗病毒和野生病毒几乎完全相同,这表明两种病毒之间存在独特的关系(Roeder 等人,2006 年),并且 K37/70 能够恢复毒力并多次重新获得在牛群中传播的能力。因此,似乎可以非常迅速地恢复毒力,而不需要积累大量的点突变。20 世纪 90 年代,在肯尼亚和坦桑尼亚,一种具有 Kabete O 基因特征的病毒似乎从出现牛瘟临床症状的牛身上分离出来。这可以表明 Plowright TCRV 具有类似的恢复毒力的潜力。可以说,TCRV 的唯一缺点是其耐热性。为了实现独立于冷链的配送系统,Mariner 等人(1990 年)通过修改干燥周期和稳定剂开发了一种更耐热的 TCRV 变体。按照新方法制成的疫苗通常被称为 Thermovax,只要避免阳光和过热,就可以在热带地区环境温度下在野外使用长达四周,无需冷链支持。恢复毒力并多次恢复在牛群中传播的能力。因此,似乎毒力可以非常迅速地恢复,而不需要积累大量的点突变。20 世纪 90 年代,肯尼亚和坦桑尼亚从出现牛瘟临床症状的牛身上分离出一种具有 Kabete O 基因特征的病毒。这可以表明 Plowright TCRV 具有类似的恢复毒力的潜力。可以说,TCRV 的唯一缺点是它的耐热性。为了实现独立于冷链的分销系统,Mariner 等人 (1990) 通过修改干燥周期和稳定剂开发了一种更耐热的 TCRV 变体。根据新方法制造的疫苗通常称为 Thermovax,可以在热带地区环境温度下在野外使用长达四周,无需冷链支持,只要避免阳光和过热即可。这恢复毒力并多次恢复在牛群中传播的能力。因此,似乎毒力可以非常迅速地恢复,而不需要积累大量的点突变。20 世纪 90 年代,肯尼亚和坦桑尼亚从出现牛瘟临床症状的牛身上分离出一种具有 Kabete O 基因特征的病毒。这可以表明 Plowright TCRV 具有类似的恢复毒力的潜力。可以说,TCRV 的唯一缺点是它的耐热性。为了实现独立于冷链的分销系统,Mariner 等人 (1990) 通过修改干燥周期和稳定剂开发了一种更耐热的 TCRV 变体。根据新方法制造的疫苗通常称为 Thermovax,可以在热带地区环境温度下在野外使用长达四周,无需冷链支持,只要避免阳光和过热即可。这

从挪威养殖大西洋比目鱼 Hippoglossus hippoglossus 细胞培养中分离诺达病毒
摘要:本文介绍了从患有病毒性脑病和视网膜病变 (VER) 的大西洋比目鱼 Hippoglossus hippoglossus 细胞培养物中分离诺达病毒的方法。细胞系 SSN-1 接种了患病幼鱼的组织材料(首次喂食后 60 天)。接种后约 5 天出现广泛的细胞病变效应 (CPE),在同一细胞系中传代数次后也观察到了这种现象。在间接免疫荧光试验中,感染培养物的细胞与抗鲈鱼 Dicentrarchus labrax 诺达病毒的抗血清表现出反应性。使用针对条纹鲈 Pseudocaranx dentex 诺达病毒的特异性引物,用逆转录聚合酶链反应 (RT-PCR) 分析感染细胞,结果得到预测大小的产物。感染的 SSN-1 细胞的电子显微照片显示病毒颗粒大约小于 30 纳米。用受感染的 SSN-1 细胞的上清液对大西洋大比目鱼幼体(孵化后 4 天)进行攻击,导致 VER 的产生,这通过对攻击后第 9 天采集的幼体样本进行的免疫组织化学检测得到证实。目前的结果表明,使用 SSN-1 细胞系分离了来自大西洋大比目鱼的诺达病毒,并且在细胞培养中繁殖的病毒保留了毒力。
人类 iPSC 亚型定向分化为心房和心室心肌细胞
15. 将 Matrigel 包被的培养板和 hiPSC 培养基预热至 20-25 C。16. 从预包被的培养板中吸出 Matrigel 并加入 hiPSC 培养基(6 孔板每孔 2 ml)。17. 将 9 ml hiPSC 培养基加入到 15 ml 离心管中。18. 将低温小瓶直接转移到 37 C 水浴中并观察解冻过程。当管中大部分内容物解冻并仅剩下一小块冰时,迅速取出并用 70% 乙醇彻底清洗。19. 小心地将细胞逐滴转移到准备好的带有培养基的 15 ml 离心管中。以 200 3 g 的速度离心 5 分钟。20. 小心吸出上清液。将沉淀物悬浮在 hiPSC 培养基(例如 1 ml)中,并接种到准备好的 Matrigel 包被的培养板上。前 24 小时加入 1 ml/ml 2 mM Thiazovivin(最终浓度 2 m M)。21. 如果 24 小时后细胞附着良好,则用 hiPSC 培养基更换培养基。如果附着力较低,再加入 1 ml/ml 2 mM Thiazovivin(最终浓度 2 m M),培养 24 小时。从第二天开始,每天更换培养基,每孔(6 孔)加入 2 ml hiPSC 培养基。继续“hiPSC 传代和维护”,步骤 1-8。

附睾基础细胞和类器官的三维细胞培养:毒理学的新工具
精子是在睾丸中形成的,但必须通过附睾过渡才能获得运动能力和受精的能力。附睾是一个单一的小管,其中包括几个在解剖学和生理上不同的基础。伪分层的上皮由多种细胞类型组成,包括主要细胞,透明细胞,狭窄细胞和顶端细胞,这些细胞与附子症的腔内齐聚。基底细胞存在于上皮的底部,其中包括巨噬细胞/单核细胞,单核吞噬细胞和T淋巴细胞的光环细胞也存在。这个综合精子成熟过程的几个方面已经建立了很好的确定,但是很多知识仍然很少。鉴于附睾的功能障碍与男性不育症有关,需要研究附睾功能的体外工具和附睾精子成熟。我们的实验室和其他人以前已经开发了人,大鼠和小鼠上皮细胞系,这些细胞系已用于解决某些问题,例如关于附睾中的junc蛋白的调节,以及北苯酚的毒性。鉴于附睾上皮包含多种细胞类型,但是,3D体外模型提供了一种更全面和现实的工具,可用于研究和阐明附子功能的多个方面。©2024作者。Wiley Perigonicals LLC发布的当前协议。本文的目的是提供有关大鼠附子基础细胞的大鼠附子器官的制备,维持,传代和免疫荧光染色的详细信息,我们已证明这是大鼠附子症中的一种成年干细胞。
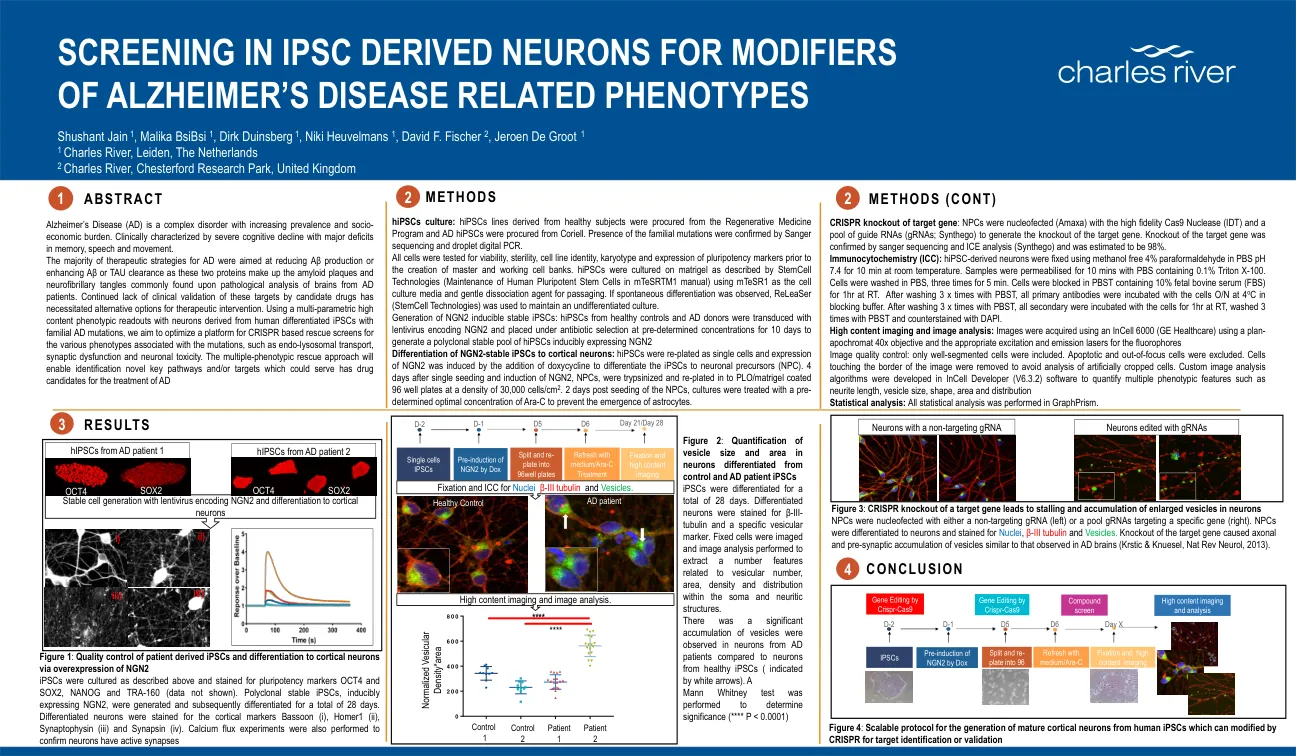
在 IPSC 衍生的神经元中筛选阿尔茨海默病相关表型的修饰因子 | Charles River
hiPSC 培养:来自健康受试者的 hiPSC 系来自再生医学计划,AD hiPSC 来自 Coriell。通过 Sanger 测序和液滴数字 PCR 确认存在家族性突变。在创建主细胞库和工作细胞库之前,对所有细胞进行了活力、无菌性、细胞系身份、核型和多能性标志物表达测试。hiPSC 在基质胶上培养,如 StemCell Technologies(mTeSRTM1 手册中的人类多能干细胞维护)所述,使用 mTeSR1 作为细胞培养基和温和的解离剂进行传代。如果观察到自发分化,则使用 ReLeaSer(StemCell Technologies)来维持未分化培养。 NGN2 诱导的稳定 iPSC 的生成:健康对照和 AD 供体的 hiPSC 被编码 NGN2 的慢病毒转导,并在预定浓度的抗生素选择下放置 10 天,以生成可诱导表达 NGN2 的 hiPSC 多克隆稳定池。NGN2 稳定的 iPSC 向皮质神经元的分化:将 hiPSC 重新接种为单细胞,并通过添加强力霉素诱导 NGN2 的表达,以将 iPSC 分化为神经元前体 (NPC)。单次接种和诱导 NGN2 4 天后,将 NPC 用胰蛋白酶消化并重新接种到 PLO/matrigel 包被的 96 孔板中,密度为 30,000 个细胞/cm 2 。在 NPC 接种 2 天后,用预定的最佳浓度的 Ara-C 处理培养物以防止星形胶质细胞的出现。

开发药物诱导耐药性肿瘤模型...
摘要 在过去的几十年里,癌症的治疗前景发生了变化。由于先进的癌症生物学、功能成像和下一代测序技术,我们对癌症及其治疗方法的理解得到了极大的提高。癌症治疗的关键挑战之一是如何有效杀死癌细胞,同时保持正常细胞完好无损。随着特定的致癌驱动因素被成功识别,靶向治疗在临床实践中取得了巨大成功。然而,获得性耐药性仍然是靶向癌症治疗持续成功的主要障碍。那些最初对靶向治疗反应良好的患者最终会对治疗产生耐药性。揭示潜在机制将有助于开发克服耐药性的新疗法。为了模拟长期药物治疗后的临床耐药性,我们通过对荷瘤小鼠或癌细胞系持续给药靶向药物建立了一组药物诱导的耐药肿瘤模型,涵盖了一系列一线靶向药物,包括 Sotorasib、Palbociclib、Ibrutinib、Capmatinib、Fulvestrant、Tamoxifen 和 T-DM1。耐药模型来自含有特定致癌驱动因素的敏感模型。经过靶向药物治疗和多次传代,耐药模型表现出稳定的耐药表型。鉴于靶基因突变、旁路激活或肿瘤微环境演变可促进治疗耐药和癌症进展,我们进行了转录本和蛋白质分析以探索分子机制,并评估了在已建立的模型中克服耐药性的潜在治疗策略。总之,药物诱导的耐药肿瘤模型为更好地理解耐药机制和加速开发下一代抗癌药物提供了有希望的机会。