XiaoMi-AI文件搜索系统
World File Search System信号肽
通过工程生物传感器
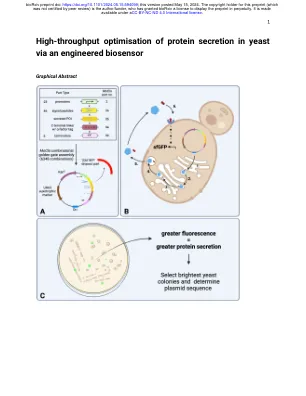
高价值蛋白质和酶的分泌是合成生物学经济的基础;允许在生产过程中连续发酵和蛋白质纯化而无需细胞裂解。大多数真核蛋白分泌由N末端信号肽编码;但是,信号肽序列变化对给定蛋白的分泌效率的强大影响尚未很好地定义。尽管天然信号肽序列多样性高,但大多数重组蛋白分泌系统仅采用少数表征良好的信号肽。此外,启动子和终止剂的选择可以显着影响分泌效率,但筛选众多遗传构建体以使最佳序列效率低下。在这里,我们调整了酵母G蛋白偶联受体生物传感器,以测量与任何感兴趣蛋白共归因于肽的肽标签的浓度。蛋白质分泌效率可以通过诱导受体激活下游的荧光报告基因来量化。此功能可以使用一锅组合金门克隆组装,对6000多个启动子,信号肽和终结器的6000多个组合进行高通量筛选。我们证明了这种生物传感器可以快速识别分泌和量化分泌水平的最佳组合。

蛋白质工程方法增强酿酒酵母真菌漆酶的产量
摘要:腐生担子菌分泌的漆酶是一种多功能生物催化剂,仅需氧气即可氧化多种芳香族化合物。酿酒酵母是真菌漆酶工程的首选宿主。为了帮助酵母分泌活性酶,天然信号肽通常被酿酒酵母α交配因子(MF α 1)的前原前导序列取代。然而,在大多数情况下,只能获得基础酶水平。在酿酒酵母中与α因子前原前导序列融合的漆酶的定向进化过程中,我们证明了信号肽中积累的突变显著提高了酶的分泌。在这里,我们描述了为增强在酿酒酵母培养物液体提取物中检测到的漆酶活性而实施的不同蛋白质工程方法。我们通过使用实验室中连续进行的漆酶进化活动获得的适应性最强的突变 α 因子前导序列,证明了天然和工程漆酶的分泌得到改善。我们还特别关注了蛋白质 N-糖基化在漆酶生产和特性中的作用,以及通过共识设计引入保守氨基酸,从而能够表达酵母原本不会产生的某些漆酶。最后,我们修改了在之前的定向进化活动中积累的漆酶编码序列 (CDS) 突变的贡献,这些突变促进了酶的生产。

烟草中重组蛋白质产生的亚细胞积累设计的进步
植物分子种植(PMF)已成为生物技术中有前途的方法。这项技术利用植物作为生物反应器的力量,将其转变为“绿色工厂”,以生产有价值的重组蛋白(例如治疗性和工业酶)。PMF利用植物的复杂生物合成机制产生复杂的重组蛋白,比确定的生产方法(如微生物发酵和哺乳动物细胞培养)具有多种优势[1,2]。例如,可以以相对较低的成本大规模种植植物,从而减少蛋白质生产的总费用。基于植物的系统可以通过增加栽培区域来轻松扩展,从而适合于高产产量。植物不含有人类病原体,从而最大程度地减少了可能影响微型和哺乳动物系统的病毒,王室或其他有害药物的污染风险。植物不会产生内毒素,这些内毒素在细菌系统中很常见,并且会使纯化过程复杂化并带来安全风险[3,4]。除了这些常见的福利外,植物还能够提供巨大的灵活性和工程潜力,从而为生产各种蛋白质的生产能够满足个性化的自定义需求。已经探索了各种植物物种,包括烟草植物[5,6],腐烂悬浮液[7-9]和水稻种子[10-13],作为生产和传递注册重组蛋白的平台。GT13A信号肽GT13A信号肽这些不同的植物生物反应器平台的每种类型都在生产效率,可容纳,可扩展性和成本效益方面具有明显的优势。例如,水稻种子具有自然促进蛋白质积累的专门储存器细胞器,在植物和后期内提供了稳定性。
工程小金属结合蛋白标签可提高大肠杆菌周质中重组人类生长激素的产量
融合蛋白在大肠杆菌重组蛋白的生产中起着重要作用。它们主要用于细胞质表达,因为它们可以设计为增加目标蛋白的溶解度,然后可以通过亲和层析轻松纯化。相反,融合蛋白通常不包含在用于周质生产的构建体设计中。相反,插入信号序列以将蛋白质转运到周质中,并添加 C 端 his-tag 以进行后续纯化。我们的研究小组提出从欧洲亚硝化单胞菌周质中分离的小金属结合蛋白 (SmbP) 作为一种新的融合蛋白,用于在大肠杆菌的细胞质或周质中表达重组蛋白。SmbP 还允许通过使用 Ni(II) 离子的固定化金属亲和层析进行纯化。最近,我们通过将 SmbP 标记蛋白的天然信号肽与取自果胶酸裂解酶 B (PelB) 的信号肽进行交换,优化了 SmbP 标记蛋白的周质生产,从而大幅增加了蛋白产量。在这项工作中,我们表达并纯化了 PelB-SmbP 标记的可溶性生物活性人类生长激素 (hGH),并获得了迄今为止报道的该蛋白的最高周质产量。在 Nb2-11 细胞上测试的其活性相当于 50 ng mL 1 的商业生长激素。因此,我们强烈建议使用 PelB-SmbP 作为蛋白标签,用于大肠杆菌周质中 hGH 或其他可能的目标蛋白的表达和纯化。

噬菌体分析:带有肽信号的内胆素和...
一些内olysin在N末端部分包含符号年度释放(SAR)域(SAR)域,首先由Xu等人描述。(2004),允许膜锚定。内ol丝蛋白含有肽切割刀信号(PS)的信号,该信号通过宿主的SEC机械(Frias; Frias; frias; helocrocristine; Ramirez,2013; Escobedo et al。,2019年)控制了内叶蛋白在periplasm中释放的信号。一些研究指出,这些结构域内的疏水程度以及信号肽(PS)的疏水程度有一定的保留。最近的研究表明,其中一些内olysin能够渗透到革兰氏阴性细菌的外膜(Lim等,2014)。因此,肽信号和SAR代表了在克细菌中外源作用的抗菌蛋白的有前途的来源

基于High-A ...
带有PCR纯化套件。此后,从含有IL-2和BI-特异性基因的预先形成的慢病毒F9质粒和BI-特异性F9质粒中扩增了IL-2信号肽和双特异性AVRAN-BIS或DURAN-BIS蛋白构建体。使用与KOD DNA聚合酶的PCR反应进行扩增,与噬菌体质粒同源的引物(补充表S1中的引物为G)。此后,使用吉布森装配方法的方案将PCR产物克隆到开放噬菌体质粒中,然后转化为电竞争性的大肠杆菌。使用Allin™Red Taq Mastermix进行了用于Gibson组装和质粒插入验证的菌落PCR,并将上述序列呈阳性的菌落转移到含有氨苄青霉素LB(25 µg/mL)和

依赖T6SS依赖性效应子RE78 ETLI MIM1有益细菌竞争
简单摘要:根瘤菌ETLI MIM1(REMIM1)具有活性在自由生活和共生中的VI型蛋白质分泌系统。T6SS是一种纳米芳烃,将称为效应子的蛋白质分泌为真核和原核靶细胞。REMIM1 T6SS基因簇编码有毒效应子(RE78)以及免疫蛋白(RE79),如在大肠杆菌中表达时所证明的。另外,观察到RE78蛋白的毒性作用在细胞质之外,因为仅当将信号肽添加到其中时才发生对大肠杆菌的毒性作用。RE79在Remim1 Periplasm中发现,并且与T6SS的易位无关。此外,RE78/RE79对还参与细菌竞争和结节占用率。更好地理解该分泌系统的作用对于选择高度竞争性根茎的接种剂可能非常有用。

使用元基因组采矿方法从高山土壤的质体中寻找新的降解酶
塑料材料,包括微塑料,即使在欧洲阿尔卑斯山等偏远和寒冷的环境中也积聚在所有类型的生态系统中。这种污染对环境和人类构成了风险,需要解决。使用大约3,000 m a.s.l.的东部瑞士阿尔卑斯山收集的土壤的shot弹枪DNA宏基因组学,我们确定了可能降解塑料的基因及其蛋白质。我们通过差异丰度分析筛选了质体和大块土壤的宏基因组,并与专门针对推定的塑料降解基因的特定数据库进行了基于相似性的筛选,并选择了具有信号肽的高概率的基因,用于信号肽的细胞外导出和高信任的功能率。此过程导致了9个候选基因的最终列表。预测蛋白的长度在425至845个氨基酸之间,预测产生这些蛋白质的属主要属于Caballeronia和Bradyrhizobium。我们使用异源表达进行了功能验证,然后进行上清液的酶测定。测试的九种蛋白质中的五种显示出我们使用酯酶测定时的活性显着增加,而从候选基因(一种水解酶型酯酶)中的五种蛋白质中的一种显然具有最高的活性,高于双倍以上。,我们仅用来自酯化酯酶测定中五个候选基因的蛋白质对塑料类型的塑料降解和生态®进行荧光测定,但是像阴性对照一样,这些蛋白质并未显示出任何偶尔的活性。相比之下,阳性对照的活性(包含文献中已知的Pla降解基因插入物)是阴性对照的20倍以上。这些发现表明,在计算机筛选中进行功能验证,适合查找新的降解酶。尽管我们只发现了一种新的酯酶酶,但我们的方法有可能应用于任何类型的土壤和各种生态系统中的塑料,以快速有效地寻找新的塑料降解酶。

纽卡斯尔病毒病毒基因型VII.1.1的分子表征来自埃及野鸭鸭,具有神经表现
在埃及,纽卡斯尔病毒病毒(NDV)的基因型VII菌株在家用水禽中是温和的,被认为是储层。这是从鸭子中检测NDV GVII.1.1的第一份报告,显示出高死亡率和神经表现的严重临床体征,此外,对全HN和F基因进行了NDV和分子表征的分离。在当前的研究中,使用针对NDV和基质基因融合基因的禽流感染基因(AIV)的融合基因(AIV)研究了16个后院野鸭群羊群,通过实时RT-PCR研究了严重的神经迹象。14只鸭羊群测试的AIV阳性,只有两只羊群对NDV感染呈阳性。ndv,然后对全Hn和F基因进行测序。F和HN基因的系统发育分析表明,这些菌株用NDV基因型VII 1.1聚集。f基因具有特定的突变,将其聚集在一个新的分支中,与疏水性含量含量重复(HRC)相比,信号肽,N30S,T324A和480K在信号肽,N30S,T324A和480K中都聚集了它们。与从同一鸭的气管中分离出的菌株相比,从大脑分离的NDV的鸭子菌株具有N294K的N294K,这可能在跨越血脑屏障中起作用。HN蛋白具有特异性突变,将它们聚集在新的分支中,其突变为A4V,R15K在细胞质区域,跨膜结构域中的A28T和HRA中的S76L。此外,HN蛋白具有A50T,S54R T232N,P392S和T443V,并且在本研究中特异性的菌株中和菌株中检测到多个突变(N120G,K284R,S521T),可以改变病毒抗原性。当前的研究表明,NDV菌株从埃及循环的基因型VII持续演变,鸭子的致病性增加。目前的发现表明,迫切需要对鸭子和鹅进行疫苗接种,并用杀死的NDV疫苗疫苗,以减少因病毒感染而导致的经济损失,并防止向鸡有助于埃及控制ND控制的鸡的传播。

抑制剂Angptl3及其对高脂血症和动脉粥样硬化的影响
和HTG导致急性胰腺炎(2)和心血管疾病(CVD)。(3,4)因此,寻求降低异常高的LDL和TG水平的靶标更有效地预防心脏病,中风和胰腺炎血管生成素样蛋白样蛋白3(ANGPTL3)是460氨基酸(AA)糖蛋白,主要由Liver分泌。angptl3包含一个N末端区域,预计本质上是无序的,一个卷曲的螺旋区域和C末端纤维蛋白原样域。(5)ANGPTL3基因的结构如图1所示。在蛋白质被裂解和糖基化后,产生了与结合和抑制脂蛋白脂肪酶(LPL)和肝脂肪酶(HL)的结合和抑制脂蛋白脂肪酶(LPL)和肝脂肪酶(HL)的N末端片段,该片段与结合和抑制脂蛋白脂肪酶(LPL)涉及。同时,分泌需要16-AA信号肽的C-末端纤维蛋白原样域(6),并参与血管生成。此功能类似于血管生蛋白的功能(7,8)