XiaoMi-AI文件搜索系统
World File Search System信息学

电气工程和信息学公告
道路缺陷检查是维持良好的运输基础设施的至关重要的任务,因为道路表面障碍会影响用户的舒适性,降低车辆零件的寿命并造成道路伤亡。近年来,由于其出色的性能和高计算能力的可用性,机器学习在包括对象检测在内的各个领域都得到了广泛的调整,这通常是其模型培训所需的。许多作品都使用基于机器学习的对象检测算法来检测缺陷,例如建筑物和道路中的裂缝。在这项工作中,使用自定义的道路裂缝和坑洼数据进行了Yolov5,Yolov6和Yolov7模型,并对其性能进行了评估和比较。数据集中的实验表明,Yolov7的性能最高,MAP@0.5得分为79.0%,推理速度为0.47 m,用于255个测试图像。

水文信息学概念
第一章确立了城市排水对人类社会的至关重要性。城市雨水和废水的管理日益被视为一项重要的业务流程,随着英国水务服务业的私有化,这种观点更加凸显。社会对行业提出的立法和监管要求,确保为客户提供高效的服务并有效保护自然环境,正在培养一种管理城市雨水和废水过程的整体观点。这种观点得到了先进建模工具的支持,这些工具使工程师能够从“源头到汇点”对综合雨水和废水管理过程进行建模,即从降雨、生活和工业废物到对受纳水体的影响。此外,对效率和效力的需求正在鼓励创新方法来减少源头流量、实时控制流量、修复现有资产并提高整个系统的性能。信息技术,即结合了计算、电信和微电子技术的技术,被水务行业视为提高效率和促进决策的一种手段。因此,水行业对信息技术的应用正在迅速增长,尽管是零碎的,而且更多的是由现有技术而非用户需求推动的。对于所谓的先进信息技术来说尤其如此,包括软件工程、人工智能和人为因素计算。用户需要一种综合的方法将水作为业务流程和所有人的资源进行管理,这需要一个新的概念框架。据称,水信息学提供了这样一个框架,其中信息技术的进步可以被利用来为那些与水生环境有关联的人们谋福利。这一说法将根据水科学家和工程师如何运用力学、地理、生物、土木工程,尤其是信息技术方面的进步来检验。

系统论、控制论和信息学
版权所有 © 2011 国际信息学和控制论研究所。保留所有权利。未经出版商书面许可,不得以任何形式或任何手段(包括复印、录制或其他方式)复制、存储本出版物的任何部分或通过任何电子或机械方式传输本出版物的任何部分。允许摘录并注明来源。图书馆可以复印用于私人用途。教师可以免费复印单独的文章用于私人用途,供非商业课堂使用。如需其他复印、重印或转载许可,请写信给 IIIC 版权经理,地址:13750 West Colonial Dr, Suite 350-408, Winter Garden, FL 34787, U.S.A. 美国印刷。ISSN:1690-4532 pp200302MI1469

地理信息学的应用
正如您现在所知,地理信息学是各种学科的协同作用,并且由于其应用主要面向自然和人为环境管理的现实问题,因此它正在迅速扩大其影响力。各种机构和组织为印度的发展做出了重大贡献。印度政府现已将地理信息技术及其衍生成果纳入其许多国家级计划中。了解它们很重要,因此,本单元与印度的地理信息学情况特别相关。在本单元中,您将学习在塑造印度当今地理信息行业方面发挥重要作用的主要机构。您还将学习涉及地理信息学的主要国家举措、参与地理信息学教育的大学和机构,这将使您对地理信息技术的范围和市场潜力以及职业前景有所了解。预期学习成果 _____________________________________

健康信息学讲座
不是会员吗?立即注册,以访问有关基础医学和临床医学的800多个教育视频。使用新视频内容免费获得每周免费更新。以明确的概念学习医学科学。主要部分包括:1。前庭眼反射2。视觉系统(有多个部分)3。diencephalon(大脑的最高部分)4。丘脑及其功能5。下丘脑及其角色6。基础核(基底神经节)及其连接7。自主神经系统(A.N.S.)肾上腺素和非肾上腺素8。动眼交感途径和霍纳综合征的解剖学9。脑皮质和语音10。睫状神经节及其连接11。pterygo-palatine Fossa和pterygopalatine神经节12。otic Ganglion,Bulbar和Pseudobulbar Palsy 13。癫痫(有多个部分)14。神经解剖学简介15。眼科神经,泪,额叶和鼻腔神经16。动眼神经及其临床相关17。颅神经(基本概念),具有骨膜神经及其临床相关性18。脑脊液(C.S.F.)和心室系统19。电动机系统和下降区域20。内耳 - 听觉系统21。神经供应泪腺22。神经系统的组织学文本还涉及与神经系统有关的各种疾病和疾病,例如阿尔茨海默氏病,老年痴呆症和其他形式的神经变性。总的来说,这个全面的概述旨在提供对神经解剖学的详细理解,涵盖理论和实际方面。一种通常与衰老有关的疾病,其特征是认知能力下降和记忆力障碍。宏观变化包括高级大脑区域的变性。显微镜,淀粉样蛋白斑块和缠结是明显的。有趣的是,已经发现大约一半的患有早期痴呆的患者在淀粉样蛋白前体蛋白基因中携带突变。这种情况与其他神经退行性疾病(例如Lou Gehrig病和多发性硬化症)具有相似之处,这些疾病会影响神经途径和可塑性。断断续续的疾病,例如幻影疼痛,也可以与中枢神经系统功能障碍有关。血管问题,包括高血压和中风,是需要考虑的其他因素。在语言动力学,学习缺陷,癫痫和精神疾病(如抑郁症和精神分裂症)的领域中表明了神经结构与功能之间的复杂相互作用。小脑功能与运动控制之间的关系也值得注意。触觉感知,音调歧视和语言能力都受到神经系统障碍的影响。必须认识到这些缺陷不是微不足道的定位,而是与更广泛的神经网络互连。
电气工程和信息学的教师
劳动力安置。决策准备技术:基于统计和启发式方法的方法:灌输管理:创新管理工具,创新管理机构,资金模型和典型应用程序的呼吁。科学研究和技术发展的组织,衍生公司的业务模型。对未来的技术构想的概念,确定技术突破的方法,对产生变化的管理。标准化过程,组织及其对技术市场的后果。创新过程中的知识产权:保护技术创造,邻近权利,保护数据库。IP权利的新趋势:免费软件许可模型。产品开发和产品介绍的过程,市场研究和营销方法。IT技术在产品和业务开发中的作用,它们对价值创造的贡献。https://portal.vik.bme.hu/kepzes/targyak/vitmak47/en/

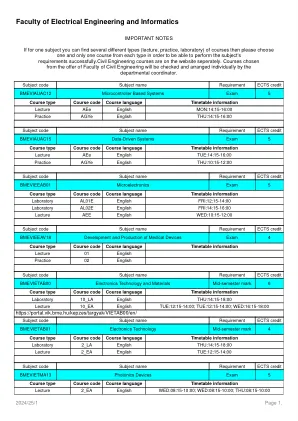
电气工程和信息学公告
由于化石燃料的使用和化石燃料的限制增加,印度能源行业正在迁移到非惯性能源资源,例如太阳能,风能生产和燃料电池。非常规能源的不可预测性使得在将电气系统合并时很难平衡,因此必须将存储设备掺入网格中。在带有风力涡轮机(WTG)和电池储能系统(BESS)的微电网系统中,BES可以在盈余生成期间保留能量,并在需求峰值的时期将其释放到网格中。建议的技术通过采用人工兔优化(ARO)算法来确定BES的收费(SOC)时间表(SOC),从而最大程度地减少了客户的能源成本。能源存储的健康状况(SOH)被纳入衰老系数,这导致贝丝(Bess)保持保守的行为以保持其寿命。使用使用时间(TOU)关税,仿真结果表明,有很大的可能性可以增加消费者在网格连接的微网格中的节省。仿真结果表明,通过有效安排BESS功率管理技术,提出的方法提高了许多分配系统功效。


电气工程和信息学公告
这项研究的目标是设计和优化光伏/风力涡轮机/电池系统。该应用是在地中海地区Bejaia(Algeria)地区进行的,该地区由于地理位置而在该地区太阳能和风能非常可利用。总入射能量方法用于开发正在考虑的设备。为了优化功率,应用模糊逻辑控制(FLC),并突出此最大功率跟踪(MPPT)策略的好处,将其与wisturb and Observe(P&O)方法进行了比较。已经应用了电源管理控制。显示和分析了三个不同日期的发现,以证明建议系统的适用性。使用Homer软件评估了检查的系统,以证明Bejaia位置的多个来源的最佳可行整合。可再生能源的增加,这是研究的主要新颖性和目标,因此PV/风系统中电池的压力较小。这是由于建议的准确尺寸程序和FLC算法。提出了在各种太阳照射和风速速度曲线下的建议研究的发现,以证明其适用性。

电气工程和信息学公告
This paper presents the application of two swarm intelligence techniques, multi-objective artificial bee colony (MOABC) and multi-objective particle swarm optimization (MOPSO), to the optimal design of a complementary metal oxide semiconductor (CMOS) low noise amplifier (LNA) cascode with inductive source degeneration.目的是在电压增益和噪声数字之间实现平衡的权衡。优化的LNA电路在2.4 GHz的运行量为1.8 V电源,并在180 nm CMOS过程中实现。在MATLAB中实现了两种优化算法,并使用ZDT1,ZDT2和ZDT3测试功能进行了评估。然后使用Advance Design System(ADS)模拟器模拟了优化的设计。结果表明,MOABC和MOPSO技术在优化LNA设计方面是实用有效的,从而比以前发表的作品更好地性能,增益为21.2 dB,噪声图为0.848 dB。