XiaoMi-AI文件搜索系统
World File Search System内源


通过内源启动子进行有效的定向诱变……
和缺失分别以+和-表示。i 使用酶StuI对T0纯合突变体进行限制性消化筛选。野生型Solanum etuberosum产生消化的PCR带(蓝色箭头),而突变植物产生对StuI消化有抗性的PCR带。J、k CR-SeSP5G突变体在短日照条件下开花,而野生型在短日照条件下不能开花。比例尺:1厘米;NF,无花。


对内源经济增长及其平衡生长路径的数值模拟
摘要在本文中,我模拟了一个经济如何内源性增长,并达到平衡的生长道路,假设家庭在MDC下的行为(最大程度的舒适度)基于基于舒适性的程序,其中MDC表示家庭对其收入和资产组合感到最舒适的状态。尽管在动态的经济增长模型中,数值不容易模拟通往稳定状态的道路,在这种模型中,家庭表现出产生理性期望的行为,但如果家庭应该在基于MDC的程序下行事以达到稳态状态,这很容易。模拟结果表明,尽管存在一些小规模效应,但经济确实可以按照理论上的预测来内源性。如果未经补偿的知识溢出受到限制,则会产生大规模的效果。较低的风险规避程度增加了增长率。此外,如果生产力相同,则经济会融合,但如果不是相同的话,则会有所不同。JEL分类:E17,E60,O11,O30,O40关键字:收敛;内源性生长;比例效应;模拟;无偿知识溢出
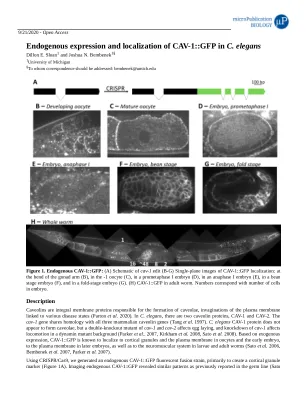
Cav-1 :: GFP在C. exkemans中的内源表达和定位
小窝蛋白是负责形成口洞的整体膜蛋白,与各种疾病状态相关的质膜的内陷(Parton等人2020)。在秀丽隐杆线虫中,有两个小窝蛋白Cav-1和cav-2。CAV-1基因与所有三个哺乳动物小窝蛋白基因共享同源性(Tang等人1997)。 秀丽隐杆线虫Cav-1蛋白似乎并不形成小窝,但是Cav-1和Cav-2的双敲击突变体会影响产卵,而Cav-1的敲低会影响动态突变体背景中的运动(Parker等人 。 2007,Kirkham等。 2008,Sato等。 2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。1997)。秀丽隐杆线虫Cav-1蛋白似乎并不形成小窝,但是Cav-1和Cav-2的双敲击突变体会影响产卵,而Cav-1的敲低会影响动态突变体背景中的运动(Parker等人。2007,Kirkham等。 2008,Sato等。 2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。2007,Kirkham等。2008,Sato等。 2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。2008,Sato等。2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。2008)。基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。2006,Bembenek等。 2007,Parker等。 2007)。2006,Bembenek等。2007,Parker等。 2007)。2007,Parker等。2007)。2007)。

解旋酶辅助连续编辑用于内源基因组的可编程诱变
人类基因组学面临的一个主要挑战是破译序列与功能之间的特定关系。然而,现有的用于在原生基因组背景下进行位点特异性超突变和进化的工具有限。在这里,我们提出了一种用于长距离、位点特异性超突变的新型可编程平台,称为解旋酶辅助连续编辑 (HACE)。HACE 利用 CRISPR-Cas9 来靶向进行性解旋酶-脱氨酶融合,该融合会在较大的 (>1000 bp) 基因组间隔内引起突变。我们应用 HACE 来识别 MEK1 中导致激酶抑制剂抗性的突变,剖析 SF3B1 依赖性错误剪接中各个变体的影响,并评估 CD69 刺激依赖性免疫增强剂中的非编码变体。HACE 提供了一种强大的工具,可用于研究编码和非编码变体、揭示组合序列与功能的关系以及发展新的生物功能。

非病毒内含子敲击素实现内源基因的简化和柔性靶向
(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此预印本版的版权持有人于2024年3月6日发布。 https://doi.org/10.1101/2024.03.05.582227 doi:Biorxiv Preprint

DNA甲基化使巨型病毒在动物亲属中反复内源化
5-甲基胞嘧啶 (5mC) 是一种广泛存在的沉默机制,可控制基因组寄生虫。在真核生物中,5mC 在寄生虫控制之外的基因调控中发挥着复杂的作用,但 5mC 也在许多谱系中丢失了。5mC 保留的原因及其基因组后果仍不太清楚。在这里,我们表明与动物密切相关的原生生物阿帕拉契变形虫具有转座子和基因体甲基化,这种模式让人联想到无脊椎动物和植物。出乎意料的是,变形虫中高甲基化的基因组区域源自病毒插入,包括数百种内源化巨型病毒,占蛋白质组的 14%。使用抑制剂和基因组分析的组合,我们证明 5mC 可以抑制这些巨型病毒插入。此外,替代的变形虫分离株显示出多态性巨型病毒插入,突显了感染、内源化和清除的动态过程。我们的结果表明,5mC 对于新获得的病毒 DNA 与真核生物基因组的受控共存至关重要,这使得变形虫成为了解真核生物 DNA 混合起源的独特模型。

定量的活细胞成像和计算建模为内源WNT/CTNNB1信号动力学
摘要Wnt/CTNNB1信号传导调节所有多细胞动物的组织发育和稳态,但是潜在的分子机制仍未完全理解。具体而言,缺少对内源性蛋白质行为的定量见解。在这里,我们结合了CRISPR/CAS9介导的基因组编辑和定量活细胞显微镜,以测量在生理和致癌条件下人类细胞中荧光标记,内源性CTNNB1的动力学,扩散特征和绝对浓度。最先进的成像表明,与途径的激活状态无关,CTNNB1的大量CTNNB1驻留在缓慢的细胞质复合物中。当Wnt/ctnnb1被激活时,这种细胞质CTNNB1复合物的大小会大大降低。基于我们的生物物理测量结果,我们构建了WNT/CTNNB1信号传导的计算模型。我们的综合实验和计算方法表明,Wnt途径激活通过三个调节节点调节不同亚细胞隔室的游离和复杂的CTNNB1的动态分布:破坏性复合物,核细胞质式穿梭和核定率。

通过AAV6介导的供体递送在人诱导的多能干细胞中增强的内源基因标记
在人类诱导的多能干细胞(HIPSC)中,系统地对内源性蛋白进行了带有荧光标记的内源性蛋白,可以观察到不同细胞态中的活细胞动力学。然而,通过CRISPR/CAS9诱导的编辑将氟化蛋白的精确插入到活细胞中依赖于同源指导的修复(HDR)。非同源末端连接(NHEJ)DNA修复途径通常胜过HDR,导致不可逆的内部和缺失(Indels)和低敲门效率。识别成功的HDR介导的标记事件是当目标基因在干细胞中未表达并且成功的标记可能不会立即观察到的靶基因是一个额外的挑战。为了解决这些挑战,我们使用了:1)在优化的感染多重性(MOI)下,与腺相关的病毒血清型6(AAV6)介导的DNA供体以最大的效率提供标签有效载荷; 2)滴定的多重CAS9:GRNA核糖核蛋白(RNP)量确保条件之间的HDR/indel频率平衡; 3)长放大液滴数字PCR(DDPCR),以测量编辑池中HDR产生的等位基因的频率; 4)同时推断CRISPR编辑(ICE)以检测并避免与Indels明显饱和(> 50%)。这些方法使我们能够确定有效,准确的编辑条件并恢复带有标记的单元,包括在干细胞中未表达的位点标记的单元。这些步骤共同使我们能够开发出有效的方法和工作流程,直接从具有最佳HDR和最小化indel频率的理想细胞池的克隆分离。使用这种方法,我们同时实现了荧光标记物和双重插入到四个基因中的基因,这些基因在差异过程中被打开,但最初在HIPSC中未表达,在HIPSC中,基于荧光基于荧光的标记细胞的直接选择是不可能的:TBR2,TBR2,TBXT,CDH2,CDH2,CDH2,CDH2,CDH2,CDH2(PROFEFFEREDIATION和MEGREDIATION和MEGREGRIATION和MEGRGIAL egratiation and Moggratial Genes and Cdhees and Cdhh5)and Cdhh5(cdh5)和CDH5(cdh)5(cdh)5(cdh)5(cdh5)。通过对各种GRNA序列和RNP浓度的系统评估,我们确定了达到高HDR频率的每个基因的条件,以38.6%的峰值达到峰值,同时还避免了与Inderels相关的条件,在这种情况下,很难在带有统一的等位基因中具有标记等位基因的克隆的隔离。过度,该方法提高了在hipscs中未表达的基因的荧光敲击的效率,对细胞过程的基于可靠的基于图像的观察,并可以恢复精确编辑的单声道和双重标记的克隆。我们将这些方法标准化,以产生有效且一般的工作流程,以将大型HDR介导的敲入引入HIPSC中。