XiaoMi-AI文件搜索系统
World File Search System分类学

宏基因组分析揭示健康母马与患有子宫内膜炎的母马子宫微生物组的新分类学差异
Albihn, A.、Båverud, V. 和 Magnusson, U. (2003)。从患有生育问题的母马体内分离的细菌的子宫微生物学和抗菌药物敏感性。《斯堪的纳维亚兽医学报》,44 (3–4),121–129。https://doi. org/10.1186/1751-0147-44-121。Ballas, P.、Reinländer, U.、Schlegl, R.、Ehling-Schulz, M.、Drillich, M. 和 Wagener, K. (2021)。患有和不患有轻度子宫内膜炎的奶牛在授精时宫内可培养需氧微生物群的特征。《动物生殖学》,159,28–34。 https://doi.org/10.1016/ j.theriogenology.2020.10.018 Bicalho, MLS、Lima, S.、Higgins, CH、Machado, VS、Lima, FS 和 Bicalho, RC (2017)。牛子宫微生物群的遗传和功能分析。第二部分:化脓性阴道分泌物与健康奶牛。乳业科学杂志,100 (5), 3863–3874。https://doi.org/10.3168/jds.2016- 12061 Clemmons, BA、Reese, ST、Dantas, FG、Franco, GA、Smith, TPL、Adeyosoye, OI、Pohler, KG 和 Myer, PR (2017)。产后哺乳奶牛的阴道和子宫细菌群落。 Frontiers in Microbiology,8,1047。https://doi.org/10.3389/fmicb.2017.01047 Díaz-Bertrana, ML、Deleuze, S.、Pitti Rios, L.、Yeste, M.、Morales Fariña, I. 和 Rivera Del Alamo, MM (2021)。野外条件下马子宫内膜炎的微生物流行率和抗菌敏感性。动物,11 (5),1476。https://doi.org/10.3390/ani11051476 Diel de Amorim, M.、Khan, FA、Chenier, TS、Scholtz, EL 和 Hayes, MA (2020)。健康母马和患有子宫内膜炎或纤维化子宫内膜退化的母马子宫冲洗液蛋白质组分析。生殖、生育力和发育,32 (6), 572–581。 https://doi.org/10.1071/RD19085 Durazzi, F.、Sala, C.、Castellani, G.、Manfreda, G.、Remondini, D. 和 De Cesare, A. (2021)。 16S rRNA 和鸟枪测序数据比较肠道微生物群的分类学特征。科学报告,11 (1), 3030。https://doi.org/10.1038/s41598-021-82726-y Frontoso, R.、De Carlo, E.、Pasolini, MP、van der Meulen, K.、Pagnini, U.、Iovane, G. 和 De Martino, L. (2008)。生育问题期间马子宫内细菌分离株及其抗菌敏感性的回顾性研究。《兽医学研究》,84 (1), 1–6。https://doi.org/10.1016/j.rvsc.2007.02.008 Heil, BA, Thompson, SK, Kearns, TA, Davolli, GM, King, G., & Sones, JL (2018). 使用多种技术对马子宫常驻微生物组进行宏遗传学表征。马兽医学杂志,66,111。https://doi.org/10.1016/j.jevs.2018.05.156 Holyoak, GR, Premathilake, HU, Lyman, CC, Sones, JL, Gunn, A., Wieneke, X., & DeSilva, U. (2022)。健康的马子宫拥有独特的核心微生物群以及丰富多样且随地理位置而变化的微生物群。科学报告,12(1), 14790。https://doi. org/10。1038/s41598-022-18971-6 Hurtgen, JP (2006). 马子宫内膜炎的发病机制与治疗:综述。《兽类生殖学》66 (3), 560–566。https://doi.org/10。1016/j.theriogenology.2006.04.006 Jianmei, C., Bo, L., Zheng, C., Huai, S., Guohong, L., & Cibin, G. (2015). 鉴定对羟基苯甲酸乙酯为产于

环境 DNA 揭示了亚马逊鱼类多样性方面因地理、环境和人为影响的不同而出现的不匹配
图 4 从最终广义相异性模型中为地理(蓝色)、环境(绿色)和人为(粉色)变量生成的 I 样条线(表 S10)。来自最终 GDM 的 I 样条线,用于(a–d)分类学 β 多样性,(e–g)分类学 β 多样性的周转成分,(h–l)功能性 β 多样性,(m–o)功能性 β 多样性的周转成分。每个函数达到的最大高度表示在所有其他变量保持不变的情况下与相关变量梯度相关的 β 多样性总量。每个函数的斜率显示 β 多样性的速率及其沿相关梯度的变化。向上,上游;由上而下,上游-下游。地理距离单位来自 NMDS 坐标。

隐性种间杂交 Lemna×... 的表征
这些微小的自由漂浮被子植物的特殊形态对浮萍科的分类学提出了挑战。尽管分子分类学有助于阐明该科的系统发育历史,但形态学数据的一些不一致导致浮萍属经常被错误分类。最近,Lemna japonica 是 Lemna minor 和 Lemna turionifera 的种间杂交种,这一发现为此类分类学问题提供了一个清晰的解释。在这里,我们证明了 L. minor 也能够与 Lemna gibba 杂交,从而在地中海地区产生一个隐秘但广泛分布的分类单元。描述了非分类单元 Lemna × mediterranea,并将其与假定的亲本种 L. minor 和 L. gibba 的克隆进行了比较。通过核和质体标记的遗传分析以及基因组大小测量表明,两种不同的细胞型(二倍体和三倍体)起源于至少两个独立的杂交事件。尽管总体相似性很高,但形态测量、生理和生化分析表明,L. × mediterranea 在大多数定性和定量特征上处于其亲本物种的中间位置,并且两种杂交细胞型也根据某些标准分开。这些数据证明,杂交和多倍化(陆生植物进化的驱动力)有助于浮萍的遗传多样性,并可能塑造了这些主要无性水生植物的系统发育历史。

生命之树Edna metabarcoding揭示了类似的分类学丰富性,但在海港和海洋储量之间存在不同的进化谱系
fi g u r e 7在分类级别的海港和储备社区的比较。位点得分(左)和物种得分(右)图代表了由空间坐标来调节的两个第一个DBRDA轴,测试了栖息地对jaccard/bray -curtis距离的影响,在汇集重复和组合所有MOTU(从MOTU级别到订单的重要性);Jaccard差异指数是在MOTU级别(a)计算的,而Bray -Curtis的差异基于MOTU的丰度用于家庭(B)和订单(C)水平。显示出促成约束轴的最高10%分类单元。中,只有那些具有高于97%的分配身份的人才能保持在MOTU级别。SMM,Saintes-Maries-de-la-Mer。

乳业科学-Tirupati
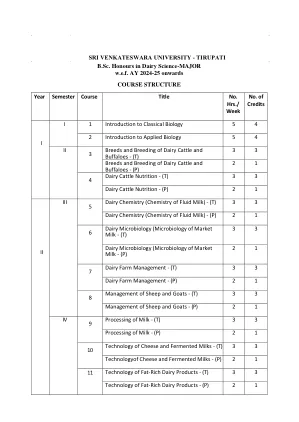
学习成果1。了解生物多样性的分类和保存原则2。了解植物的解剖,生理和生殖过程。3。关于动物分类,生理学,胚胎发展及其经济重要性的知识。4。概述细胞成分,细胞过程,例如细胞分裂,遗传和分子过程。5。理解塑造和驱动大分子和生命过程的化学原理。单元1:系统学,分类学和生态学概论1.1。系统学 - 定义和概念,分类学 - 定义和层次结构。1.2。命名法 - ICBN和ICZN,二项式和三项术命名。1.3。生态学 - 生态系统,生物多样性和保护的概念。1.4。污染和气候变化。单位2:植物学2.1的必需品。植物王国的分类。2.2。植物生理过程(光合作用,呼吸,蒸腾,植物激素)。2.3。花的结构 - 微观和宏观生成,授粉,施肥和

招聘广告
在植物的进化和系统学的博士后研究人员尽快开始。我们最初提供三年的全职职位(每周40小时)。可以再进行三年的扩展。植物的一组综合分类学(Marc Appelhans教授)正在我们新成立的团队中寻找一个积极进取的博士后。我们正在寻找一个专注于系统植物学的人,并具有下一代测序方法和生物信息学技能的经验。我们目前的研究集中在(亚)热带五经和原始科的系统基因学,分类学和生物地理学上,我们希望积极参与现有和未来的项目以及(共同)对学生项目的监督和学生助理的指导。我们鼓励自己的想法,我们希望您获得自己的项目并建立独立的研究简介。该职位提供了获得学历(Habilitation)的机会。您的职责:

咨询-II:使用对区域敏感的哈希
微生物群落的宏基因组测序产生了来自未知的微生物的简短DNA读数(Handelsman,2004),导致需要基于参考数据集的分类学识别。一种方法是从分类学上识别读取并总结结果以获得样本的分类学概况,显示了分类群体的相对丰度。但是,尽管有成熟的读取分类和分析工具的可用性,但基准测试揭示了现有方法的准确性的主要差距(McIntyre等人。,2017年; Meyer等人。,2019年; Sczyrba等。,2017年; Ye等。,2019年)。精确的识别通常会受到查询的新颖性与全基因组参考数据集和模棱两可的匹配的阻碍。此外,对大量基因组进行搜索是计算要求的。分类学识别方法采用各种策略,包括K -Mer匹配(Ames等人,2013年; Ounit等。,2015年;伍德等。,2019年; Lau等。,2019年; Lu等。,2017年),阅读映射(Zhu等人,2022),基于标记的对准(Liu等人。,2011年;米兰等。,2019年; Segata等。,2012年; Sunagawa等。,2013年)和系统发育放置(Asnicar等人。,2020年; Shah等。,2021; Truong等。,2015年)。无论如何,它们本质上都搜索了样本中的读数和参考集之间的匹配。,2017年),尤其是在众所周知的微生物栖息地(如海水或土壤)中(Pachiadaki等人。,2019年)。挑战是地球微生物多样性的很大一部分缺乏参考数据集中的近距离代表(Choi等人因此,大多数方法

根据分类隶属关系估算共识蛋白质组和代谢功能
摘要目的:元编码和元基因组测序使高度多样化的环境社区的表征能够表征。估计这些社区执行的代谢功能的挑战导致了几种最新方法的发展,其中大多数方法是根据特定基因标记量身定制的。但是,测序技术进步所产生的方法的增加,驱动了能够处理异质微生物社区数据的方法。预测通常取决于其内部分析管道,并受到基础数据库的影响,这些数据库将标记基因与特定功能注释联系起来。这限制了用户通过追踪内部数据和流程来评估预测质量的能力。最后,用户受这些方法提供的特定注释的约束(例如ec数),限制了他们根据中间结果进行进一步专业分析的能力。方法:ESMECATA预测分类学官员的共识蛋白质组及其相关功能。Esmecata的关键特征是其解释性和功能。为了支持异质测序数据的灵活整合,Esmecata利用了通过分析不同测序数据集的分析获得的分类学官方。为了深入了解每种分类学的知识并解释预测功能的相关性,Esmecata确定了在UNIPROT数据库中有记录的蛋白质组织能够辅助代表的给定官能中的分类学等级。根据阈值,将Uniprot蛋白质组的蛋白聚类并过滤,以创建共识蛋白质组。这些共有的蛋白质组会自动用功能信息(例如,EC数字,GO术语)注释,但它们也被设计为用于进一步的自定义注释工作流。功能注释在功能表中报道,该功能表可以充满分类的丰度,以产生全面的功能性文件。结果:ESMECATA预测已使用多个数据集验证,并将其与最新方法进行了比较。此外,它被应用于甲烷剂反应器的新型元编码数据集,表征了微生物群落和沼气在不同的时间点和进气条件上的产生。我们的结果证明了沼气之间的联系

整合分类学,遗传和生态数据,以探索Culuccia Peninsula(意大利NE Sardinia)的野生蜜蜂(hymenoptera,apoidea,anthophila)的物种丰富度
抽象的野生蜜蜂是本地和栽培植物的基本传粉媒介,但其种群在全球范围内正在下降。保护工作受到数据不足的阻碍,尤其是在地中海盆地中,该盆地拥有世界上一些最多样化的传粉媒介社区。尤其是在地中海最大的岛屿撒丁岛,关于蜜蜂动物区系的信息仍然有限。这项工作的目的是通过结合传统(基于形态的)分类法和DNA条形码,从东北萨尔迪亚(意大利)中未开发的半岛提供了apoidea anthophila的第一个清单。此外,还提供了鲜花的记录并在访客网络中显示,以丰富有关地中海地区野生蜜蜂与植物之间关联的稀缺数据。蜜蜂从2022 - 2023年4月至2023年10月进行采样。DNA以扩增线粒体基因cyotochrome氧化酶I的序列,然后将其与使用鉴定工具的BOLD进行比较,并通过构造邻居加入的系统发育树。收集并鉴定出属于29属的76种不同的物种和六个家族。对于61种不同的物种,总共获得了212个COI序列,其中许多物种尚未从意大利人群中测序。收集的五个分类单元是萨尔多 - 科尔斯裔人物,而六种是从撒丁岛新记录的。最后,我们重点介绍了潜在的分类问题和新的鲜花访问记录,强调需要进一步研究,以更好地了解这种多样化的昆虫的分类学和生态,以保护其保护。