XiaoMi-AI文件搜索系统
World File Search System动物细胞
基因转移到动物细胞
动物细胞的基因转移方法首次在1960年代初开发,当时使用培养的哺乳动物细胞的研究人员寻求方法来提高病毒DNA和RNA感染的效率。从那时起,引入外源遗传物质的技术已经显着多样化和改善,现在它们基于当前的分子和细胞生物学研究的大部分研究,并构成了生物技术的许多应用方面的基础。已经描述了各种方法用于基因转移到动物细胞中,包括化学和物理递送技术,使用病毒载体转移基因转移以及最近使用细菌载体转移基因转移。这些技术不仅用于在动物细胞中添加新的DNA,还构成了允许随机或靶向基因破坏和其他形式的基因组修饰的方法的基础。在过去几年中,随着研究人员已转向RNA干扰,将RNA传递到动物细胞中变得越来越重要。


哺乳动物细胞中的 CRISPR-Cas9 编辑
其他考虑因素!!• 贴壁细胞还是悬浮细胞?:两种方法都行,但悬浮细胞通常更容易(尤其是大规模培养)• 我的细胞系有核型分析数据吗?(细胞系可以是非整倍体 -> 更多等位基因可 KO)• 您所需的细胞系是否表达您需要的途径?

从细菌,植物和动物细胞瞄准的DNA
原理:UV吸收光谱基于以下原理:核酸(DNA和RNA)在特定波长(主要是260 nm)上吸收紫外线。吸收的紫外线量与样品中存在的核酸的浓度成正比。在260 nm处的吸光度与在280 nm处的吸光度比表明核酸的纯度,较高的比例表明纯核酸(蛋白质污染较少)。

哺乳动物细胞中的耦合基因编辑
通过转染短单链寡脱氧核糖核苷酸(SSODN),可以将小基因组改变引入高精度的哺乳动物细胞中。ssodns在DNA复制过程中集成到基因组中,但是由DNA不匹配修复(MMR)易于检测所得的杂化,从而阻止了有效的基因修饰。我们以前已经证明,当Ssodn中的核苷酸不匹配是锁定的核酸(LNA)时,可以避免MMR的抑制作用。在这里,我们揭示了LNA修饰的SSODN(LMOS)并未作为哺乳动物细胞中的完整实体整合,而是在靶杂交之前和之后被严重截断。我们发现,LMO的5'-arm臂中的单个额外(非LNA修饰)突变影响靶向效率,并激活了MMR途径。相比之下,3'-ARM中的其他突变不会影响靶向效率,并且不受MMR的影响。甚至更引人注目的是,3'-arr中的同源性在很大程度上是有效靶向的,暗示了大量的3'末端修剪。我们提出了一个在包括LMO降解的哺乳动物细胞中LMO指导基因修饰的精制模型。

第三单元 - 微生物,动植物,动物细胞,器官培养...
Gottlieb Haberlandt是奥地利植物学家。他是欧洲“大豆”先驱教授弗里德里希·J·哈伯兰特的儿子。Haberlandt首先指出了孤立组织和植物组织培养的可能性。他提出了通过组织培养的单个细胞的潜力,还提出了组织的相互影响可以通过这种方法来确定。哈伯兰特(Haberlandt)针对组织和细胞培养的原始断言方法已经实现,从而导致了生物学和医学的重要发现。他在1902年提出的最初想法被称为Totipentiality:“从理论上讲,所有植物细胞都能够产生完整的植物。”Gottlieb Haberlandt在1904年给出了Kranz(德语)解剖学一词,以描述陆地植物中更高效的C4光合作用中发现的专门叶片解剖结构。

从哺乳动物细胞和固定组织中分离基因组DNA从哺乳动物细胞和固定组织中分离基因组DNA
获得用于遗传研究的DNA已成为几乎所有生物医学研究的重要组成部分。本单元为全血,人体组织,培养细胞和唾液/颊拭子的简单,成本效益的DNA制备提供了更新。使用新鲜或冷冻样品时,可以常规地分离出适合下游测定法的高分子重量DNA,例如基因分型阵列或下一代测序。DNA也可以从福尔马林固定的石蜡装置(FFPE)组织中分离出来,尽管它更具挑战性,并且在下游应用中存在局限性,该DNA适合使用。在过去的几十年中,商业套件的可用性和成本降低使它们在该领域中的使用引起了人们的关注。有许多提供类似类型的提取套件的供应商,这是从人类标本中获得高质量DNA的一种经济高效的选择。大多数商业上可用的提取方法不再需要使用有毒化学物质,例如苯酚和氯仿,也使它们成为研究人员的更安全选择。商用套件通常在DNA的产量和质量方面具有手动方法(Chacon-Cortes&Griffiths,2014; Guha等,2018)。基本协议1描述
硫化氢在哺乳动物细胞,组织和器官中的生理作用
H 2 S现在被认为是多种哺乳动物细胞和组织中的内源性生理调节剂。Produced, in a regulated and cell type-dependent manner, by three major enzyme systems, cystathionine c -lyase (CSE), cystathio- nine b -synthase (CBS), and 3-mercaptopyruvate sulfurtransferase (3-MST), H 2 S is present intra- and extracellularly and interacts with proteins, DNA, and other members of the reactive species interactome (例如,氧和氮衍生的氧化剂和自由基)并在各种目标和途径上发挥作用。H 2 S的生理作用在基因转录和翻译,细胞生物能学和代谢,血管张力和免疫功能中的调节中得到充分认识,在中枢神经系统和周围神经系统的各种功能以及与生理学家和临床医生相关的许多其他领域的调节中。本综述对H 2 S在哺乳动物细胞和器官中的生理调节作用进行了全面概述。在生理状况下对这些作用的理解以及对H 2 S稳态的扰动的日益了解(例如,血管疾病,血管疾病,代谢性疾病,各种形式的中枢神经系统疾病,各种形式的中枢神经系统疾病,对跨性别疾病的疾病,其他机构的疾病以及其他机理疗法的诊断和诊断的新机会。在这种情况下,基于H 2 s的替换(通过H 2 s-释放的小分子)的新型实验治疗方法已经出现,并正在转化为临床竞技场。在本综述中突出显示,由于生物合成和/或降解增加,在某些疾病中,H 2 S水平在病理上降低了(例如,再灌注损伤,动脉粥样硬化,动脉粥样硬化以及许多其他形式的血管疾病,以及衰减)。在其他疾病(例如,各种形式的炎症,唐氏综合症和癌症)中,H 2 S水平增加,并且抑制H 2 S产生酶正在作为一种实验性治疗方法出现。进一步了解H 2 S的生理调节作用,再加上旨在调节H 2 S稳态的小分子的药理学和翻译科学的进步,预计将来会产生新颖的诊断和临床疗法方法。

光介导控制哺乳动物细胞中的基因表达
可以使用不同颜色的荧光蛋白同时对基因进行成像(Miyawaki,2011;Han et al.,2019)。由于分子成像探针的发展取得了最新进展,可以获得不同细胞和组织状态下的细胞基因表达模式信息(Sakaue-Sawano et al.,2008;Kohl et al.,2014;Lin and Schnitzer,2016;Sakaguchi et al.,2018)。除了荧光蛋白成像外,生物发光成像也有助于定量分析基因表达动态(Shimojo 等,2008;Imayoshi 等,2013;Imayoshi and Kageyama,2014;Isomura 等,2017;Suzuki and Nagai,2017;Sueda 等,2019)。尽管生物发光探针的多色成像最初在技术上受到限制,但最近开发的短波长和长波长荧光素酶
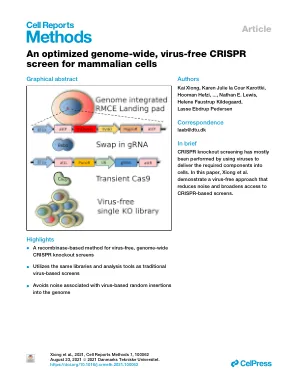
针对哺乳动物细胞进行优化的全基因组无病毒 CRISPR 筛选
Kai Xiong, 1 Karen Julie la Cour Karottki, 1 Hooman Hefzi, 2,5 Songyuan Li, 1 Lise Marie Grav, 1 Shangzhong Li, 2,6 Philipp Spahn, 2,5 Jae Seong Lee, 3 Ildze Ventina, 1 Gyun Min Lee, 1,4 Nathan E. Lewis, 2,5,6 Helene Faustrup Kildegaard, 1 和 Lasse Ebdrup Pedersen 1,7,8,* 1 丹麦技术大学诺和诺德基金会生物可持续性中心,丹麦灵比 2 加州大学圣地亚哥分校诺和诺德基金会生物可持续性中心,美国加利福尼亚州拉霍亚 3 亚洲大学分子科学与技术系,韩国水原 16499 4 韩国科学技术研究院生物科学系,大田 5加州大学圣地亚哥分校儿科,美国加利福尼亚州拉霍亚 6 加州大学圣地亚哥分校生物工程系,美国加利福尼亚州拉霍亚 7 丹麦技术大学生物工程系,丹麦林比 8 主要联系人 *通信地址:laeb@dtu.dk https://doi.org/10.1016/j.crmeth.2021.100062