XiaoMi-AI文件搜索系统
World File Search System响应元件

再生响应元件Careg监测MULLER GLIA在MNU诱导的斑马鱼视网膜损伤后的激活
fi g u r e 4皮肤DEG具有比皮肤DSG更大的基因共表达连通性,但DSGS的表达更高。(a)小提琴图显示了转录组,DSG和DEG中所有基因的总连通性(ktotal)值的分布。(b)小提琴图显示了转录组,DSG和DEG中所有基因的基因表达值的分布。表达值以每百万(TPM)的转录本为标准化。为了视觉清晰度,在“所有基因”类别中的表达值超过200 tpm的635个异常值不包括在图中。两者均为DSG和DEG的六个基因均不包括在任何分析中。在这两个图中,中间的白色钻石代表分布的中值,每个成对比较的置换测试的结果均显示为星号(*p <.05; *** p <.001; **** p <.0001)或ns(不重要)。

det mediske fakultetordinærekann,模量1,第2块
基因的启动子/增强子区域。当Carine受体结合与响应项的结合通过其配体的结合激活时,其启动子/增强子中具有响应元件的基因会在转录上向上调节。即使使用其配体激活核心受体,也不会调节其促进中没有这种DNA序列/反应元件的基因。激活核心受体的配体的例子是类固醇激素和各种营养素(脂肪酸,维生素A,维生素D)。预计不会提到下面提到的细节,但是应该说明一个短的DNA序列重复两次会导致不同的响应元素。[核心受体结合的响应元件基于两个六核苷酸(AGGCA)的重复序列。是什么赋予独特的绑定座椅
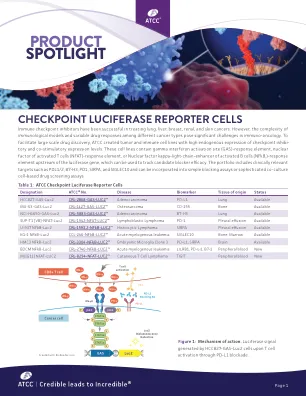
检查点荧光素酶报告基细胞
免疫检查点抑制剂已成功治疗肺,肝,乳腺癌,肾脏和皮肤癌。然而,不同癌症类型之间免疫模型和可变药物反应的复杂性在免疫肿瘤学中构成了重大挑战。为了促进大规模的药物发现,ATCC创建了具有高内源性表达的肿瘤和免疫细胞系的检查点抑制性疗程和共刺激性表达水平。这些细胞系包含伽马干扰素激活位点(气)回应元件,活化T细胞的核因子(NFAT) - 回答元件或活化B细胞的核因子Kappa-Light-chain-Enhancer(NFKB,) - 可用于跟踪候选候选者的Luciferase Gene上游的响应元件。投资组合包括临床相关

剖析p63和p53
摘要转录因子p53是最著名的肿瘤抑制剂,但其同胞p63是表皮发育的主要调节剂,也是鳞状细胞癌(SCC)中的关键致癌驱动器。尽管有多种基因表达研究,但报告的p63依赖性基因的重叠有限,因此很难破译p63基因调节网络。尤其是在研究中对p63响应元件的分析有很大不同。为了解决这种复杂的数据情况,我们提供了一个综合资源,该资源能够评估对任何感兴趣的人类基因的p63依赖性调节。我们结合广泛的CHIP-SEQ数据结合使用了一种新型的迭代基序搜索方法,以实现p53和p63结合位点,识别基序和潜在的共同因素之间的精确全球区别。我们将这些数据与增强子:基因关联整合在一起,以预测p63靶基因,并确定代表预后和治疗干预候选者的SCC中通常取消调节的基因。

香己酸通过激活PPAR-α
摘要 - 过氧化物酶体增殖物激活受体(PPAR)-α是皮肤炎症性疾病,高增生和异常分化的皮肤条件的关键调节剂。对表皮分化和皮肤屏障改善需要新的对PPAR-α激活剂的搜索。香己酸。香己酸是一种在柠檬草和柑橘类水果精油中发现的无环单丙烯羧酸。香己酸增强了PPAR响应元件(PPRE)和晶状膜形成(CE)形成的转移活性,并降低了炎性细胞因子和抗微生物肽的表达。香己酸还促进了依赖蛋白的蛋白质表达,作为CE的成分和角质形成细胞分化的标志物,以及透明质酸(HA)的合成,透明质酸(HA),一种保湿成分。这些结果表明,香氯酸可能是改善表皮屏障功能的合适皮肤治疗方法。关键词 - 香己酸,过氧化物酶体增殖物激活受体,透明质酸,促炎细胞因子,抗微生物肽

课程Vitae Carlos Antonio Molina-Nazario
教育:本科1981- 1985年(Magna cum Laude)波多黎各科学学院生物学系,里奥·皮埃尔斯·波多黎各专业:生物学小学:化学毕业生和专业专业1986- 1989年M.S.(柯蒂斯·阿什德尔(Curtis Ashendel)顾问)药物化学系,印第安纳州西拉斐特大学药学院,大专业:药物化学小学:癌症生物学1989-1991博士学位。 (Advisor, Curtis Ashendel) Medicinal Chemistry Dept., School of Pharmacy Purdue University, West Lafayette, Indiana Major: Biochemistry Minor: Cancer Biology Postdoctoral Training 1992-1994 Post-Doctoral Fellow with Dr. Paolo Sassone-Corsi CNRS-LGME, Institut De Chimie Biologique Faculté De Medicine, University Louis Pasteur, Strasbourg, France The环形响应元件调节剂(CREM)在信号转导过程中的作用。大学任命:1994-1996,妇产科助理教授(联合任期)助理教授Umdnj-new泽西医学院,纽瓦克NJ NJ 1996-2007助理教授

癌细胞国际诱导基因表达
图1创建合成cAMP响应元件结合蛋白(CREB)响应启动子。(a)腺苷信号传导的描述。腺苷(红色球)结合腺苷受体A2AR/A2BR,该腺苷受体动员相关的G蛋白(绿色)激活腺苷酸环化酶(橙色受体),并将ATP转化为3'5'- 5'-循环腺苷单磷酸腺苷(Camp)。另外,福斯科蛋白(橙色球)可以直接激活腺苷循环酶。CAMP结合蛋白激酶A(PKA)与磷酸化的CREB,该CREB结合了Plindromic DNA基序“ TGACGTCA”,激活了基因表达。(b)启动子设计和筛选示意图。cAMP响应元件基序(CRE,突出显示的黄色)被克隆在3倍重复中,两侧是鸟嘌呤“ G”(带下划线),六个散布的填充核苷酸(N)。3x Cres(灰色正方形)放在核心启动子(蓝色箭头)上游的1-6个重复中。用高斯荧光素酶(GLUC)或绿色荧光蛋白(EGFP)定量启动子活性。(c,d)HEK293T细胞在96个井板中用指示的构建体(x轴)反向转染。转染后48小时,用车辆(DMSO,浅蓝色条)或20μm福斯科林(FSK,深蓝色条)将细胞介质更改为培养基。八个小时后,对培养基进行了采样并测试了GLUC活性(RLU)。条表示n = 3实验重复的平均值,误差线代表标准误差(SEM)。**通过方差分析(ANOVA)Tukey检验,与所有其他样本相比,表示P <0.01。(E,F)流式细胞仪启动子诱导。HEK293T细胞用96个井板中的指定构建体(x轴)反向转染。转染后48小时,细胞培养基被更改为未处理的培养基(浅蓝色条),或补充了0.750 m m m腺苷(ADO,深蓝色条)的培养基。八个小时后,将细胞胰蛋白酶胰蛋白酶进行胰蛋白酶,并将其重悬于FACS缓冲液中以进行流式细胞仪。y轴表示正向散射(FSC)单元的EGFP中位荧光强度。条代表n = 3实验重复的平均值,误差线代表SEM。(g)启动子对腺苷的剂量反应性。HEK293T细胞在96个井板上反向转染,并在传说中指示的构造,然后培养48小时。然后更改培养基以添加不同的腺苷浓度,在8小时后进行采样,并测试了GLUC活性(RLU)。**通过12倍-CRE_YB的ANOVA TUKEY测试代表P <0.01,与1 m m的所有其他样品相比。每个点表示n = 3实验重复的平均值,误差线为SEM。
DNA元件系绳典型Polycomb抑制复合物1 to Human Genes
多细胞动物的摘要需要polycomb组蛋白的表观遗传抑制。后者在多种亚基X es中组装,其中两种,poly comb r ePressiv e comple x 1(pr c1)和poly comb r e:re atressiv e comple x 2(prc2),起作用,以抑制k e y de v e v elopmental基因。ho w pr c1和pr c2识别特定基因仍然是一个悬而未决的问题。在这里,我们报告了数百个DNA元素的鉴定,这些DNA元素将规范PRC1绑定到人类发育基因。我们使用该术语系列来描述在某些基因组部位在某些基因组部位显着存在的过程,尽管该复合物不太可能直接与DNA相互作用。详细的分析表明,与PRC1束缚相关的序列特征与F a v我们的PR C2结合的序列特征不同。t hrought the Genome,两种序列的特征是不同比例混合的,以产生一系列的DNA元素,这些元素的范围从主要是prc1或prc2到能够束缚这两种复合物的元素。新兴图片类似于果蝇的多梳子响应元件(PRES)对polycomb络合物的范式靶向,但可塑性是较高的。

具有改进的耦合效率的工程Luminopsins
在行为实验动物中对神经元活性的操纵对于阐明脑功能的神经元网络至关重要。光遗传学1和化学遗传学2方法对于确定遗传定义的神经元种群对电路和行为输出的贡献仍然非常有价值。两种方法都具有明显的优势,并在精确的时间尺度上对神经元亚群的活性进行了光遗传控制,并且对整个神经元群体活性的化学遗传控制较慢。以前的工作已经开发了一种工具集,该工具集通过将光发射荧光素酶融合到光遗传学的光响应元件中,从而积分光学和化学遗传学方法,从而产生发光的Opsin或Luminopsin(LMO)(LMO)3 - 5 [图。1(a)]。通过荧光素酶氧化可扩散的荧光素底物产生的生物发光会激活附近的蛋白蛋白。取决于OPSIN的生物物质特性,荧光素酶产生的光可以激发或抑制表达LMO的靶神经元。将光学和化学方法的这种整合允许在同一实验动物中同一神经元的一系列空间和时间尺度上操纵神经活动。例如,可以将整个神经元群体激活的行为成分的贡献与同一神经元子集的群体进行比较,从而通过生物发光或光遗传纤维通过光纤维在化学上激活OPSIN化学。6

全基因组识别和硅内表达...
抽象类胡萝卜素裂解酶(CCOS)酶通过产生多种杀伤型及其衍生物,在植物生长和发育中起重要作用。这些化合物对于为花朵和水果以及合成的植物激素(例如脱甲酸和strigolactone)而言至关重要。尽管其重要性,但尚未确定向日葵中CCO酶的基因家族响应。在这项研究中,我们确定了葵花籽植物的CCO基因,以填补这一知识空白。系统发育和同步分析表明,在不同植物物种中保守的Helianthus annnus cco(Hacco)基因可以根据其保守域将其分为三个亚组。使用模因工具和多个序列比对分析在HACCO基因序列中鉴定出保守的基序。顺式调节元素(CRE)分析Hacco基因表明存在与植物激素,发育以及对生物和非生物胁迫的反应有关的各种响应元件。这意味着这些基因可能会对植物激素,发育提示和干旱胁迫反应,从而在发育更具耐药作物的发育中提供了潜在的应用。属于9-CIS-环氧类胡萝卜素双加氧酶(NCED)亚组主要表现出叶绿体定位,而在其他组中发现的基因主要位于细胞质中。通过60个miRNA调节了这21个鉴定出的Haccos,表明microRNA在向日葵中基因调节中的关键作用。在干旱胁迫下的基因表达分析显示,Hanced16和Hanced19的显着上调,这是ABA激素生物合成中关键的基因。在器官特异性基因表达分析中,HACCD12和HACCD20基因在叶片中表现出较高的活性,表明在叶子色素沉着中具有潜在的作用。这项研究为未来的研究及向日葵基因家族的调节和功能的研究奠定了基础。有可能开发可用于在育种计划中使用的分子标记物,以创建对生物和非生物胁迫具有抗性的新向日葵线。