XiaoMi-AI文件搜索系统
World File Search System定向诱变

更广阔视角下的生物技术 |报告
欧盟生物技术立法(转基因立法)的现代化 欧盟委员会目前正在制定一项政策倡议,以使有关新型遗传育种技术(NGT)的立法现代化。这特别涉及针对同源基因(将基因从一个物种的一个植物转移到另一个植物)和定向诱变(在不添加新基因的情况下修改基因组上选定的位置,例如“打开或关闭”基因)可能制定的新立法。目前,此类技术仍属于现有转基因法规的管辖范围。 IUCN-NL、自然与环境(N&M)和北荷兰自然与环境联合会(MNH)赞同欧盟委员会的目标,认为调查立法是否可以现代化是一个好主意。在此过程中,N&M 和 MNH 向欧盟委员会提供了他们认为至关重要的以下原则。 IUCN-NL作为IUCN国际的一部分,对生物技术主题没有官方立场。

基于 CRISPR-Cas9 的毛霉菌诱变......
摘要:Lichtheimia corymbifera 被认为是最常见的毛霉菌之一。由于缺乏有效的基因操作工具,我们无法表征这种机会性致病真菌的致病机制和毒力因子。尽管此类技术已用于某些物种,但在毛霉目真菌中,进行定向诱变和构建稳定转化体仍然是一个巨大的挑战。在本研究中,应用无质粒 CRISPR-Cas9 系统对 L. corymbifera 进行定向基因破坏。所述方法基于 Cas9 酶引起的双链断裂的非同源末端连接修复。利用该方法,可以在乳清苷 5′-磷酸脱羧酶基因 (pyrG) 中诱导一到五个核苷酸长的短靶向缺失,从而构建尿嘧啶营养缺陷型菌株。这些菌株可作为未来基因操作研究中的受体菌株。据我们所知,这是这种临床相关真菌的首次基因改造。

基于 CRISPR-Cas9 的毛霉菌诱变......
摘要:Lichtheimia corymbifera 被认为是最常见的毛霉菌之一。缺乏有效的基因操作工具阻碍了对这种机会性致病真菌的致病机制和毒力因子的鉴定。尽管此类技术已在某些物种中得到描述,但在毛霉目真菌中,进行定向诱变和构建稳定的转化子仍然是一个巨大的挑战。在本研究中,应用无质粒的 CRISPR-Cas9 系统对 L. corymbifera 进行定向基因破坏。所述方法基于 Cas9 酶引起的双链断裂的非同源末端连接修复。利用该方法,可以在乳清苷 5'-磷酸脱羧酶基因 (pyrG) 中诱导一到五个核苷酸长的短靶向缺失,从而构建尿嘧啶营养缺陷型菌株。这些菌株可作为未来基因操作研究中的受体菌株。据我们所知,这是这种临床相关真菌的首次基因改造。
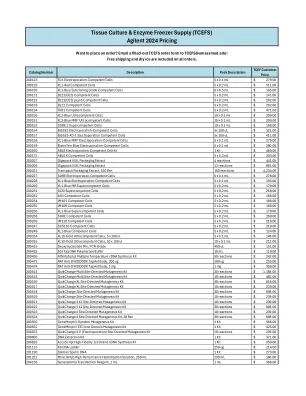
安捷伦 2024 定价
价格 200123 TG1 电穿孔感受态细胞 5 x 0.1 毫升 279.00 美元 200129 XL1-红色感受态细胞 5 x 0.2 毫升 511.00 美元 200130 XL1-蓝色亚克隆级感受态细胞 8 x 0.5 毫升 105.00 美元 200131 BL21(DE3) 感受态细胞 5 x 0.2 毫升 141.00 美元 200132 BL21(DE3) pLysS 感受态细胞 5 x 0.2 毫升 202.00 美元 200133 BL21 感受态细胞 5 x 0.2 毫升 202.00 美元 200134 TKB1 感受态细胞 5 x 0.2 毫升 471.00 美元200150 XL2-Blue 超感受态细胞 10 x 0.1 毫升 209.00 美元 200151 XL2-Blue MRF' 超感受态细胞 10 x 0.1 毫升 299.00 美元 200152 SURE 2 超感受态细胞 10 x 0.1 毫升 188.00 美元 200154 BJ5183 电穿孔感受态细胞 5x 100 微升 321.00 美元 200157 BJ5183-AD-1 电穿孔感受态细胞 5x 100 微升 411.00 美元 200158 XL1-Blue MRF' 电穿孔感受态细胞 5 x 0.1 毫升 279.00 美元 200159 EletroTen-Blue电穿孔感受态细胞 5 x 0.1 毫升 280.00 美元 200160 ABLE 电穿孔感受态细胞试剂盒 1 套 480.00 美元 200172 ABLE K 感受态细胞 5 x 0.2 毫升 303.00 美元 200207 Gigapack III XL 包装提取物 4 次反应 441.00 美元 200209 Gigapack III XL 包装提取物 11 次反应 881.00 美元 200221 Transpack 包装提取物,100 次反应 100 次反应 4,234.00 美元 200227 SURE 电穿孔感受态细胞 5 x 0.1 毫升 278.00 美元 200228 XL1-Blue 电穿孔感受态细胞 5 x 0.1 毫升 195.00 美元200229 XL1-Blue MR 超级感受态细胞 5 x 0.2 毫升 179.00 美元 200231 SCSI 超级感受态细胞 5 x 0.2 毫升 258.00 美元 200232 AG1 感受态细胞 5 x 0.2 毫升 186.00 美元 200234 JM101 感受态细胞 5 x 0.2 毫升 186.00 美元 200235 JM109 感受态细胞 5 x 0.2 毫升 186.00 美元 200236 XL1-Blue 超级感受态细胞 5 x 0.2 毫升 179.00 美元 200238 SURE 感受态细胞 5 x 0.2 毫升 269.00 美元 200239 JM110 感受态细胞5 x 0.2 mL 219.00 $ 200247 SCS110 感受态细胞 5 x 0.2 mL 219.00 $ 200249 XL1-Blue 感受态细胞 5 x 0.2 mL 124.00 $ 200314 XL10-Gold 超感受态细胞,5 x 100 ul 5 x 0.1 mL 148.00 $ 200315 XL10-Gold 超感受态细胞,10 x 100 ul 10 x 0.1 mL 212.00 $ 200415 脱氧核苷酸混合物,PCR 级 400 ul 151.00 $ 200435 10x Taq DNA 聚合酶缓冲液 10 mL 119.00 $ 200436 AffinityScript 多温度 cDNA 合成试剂盒 50 次反应 242.00 美元 200473 RAT 抗 DYKDDDDK 标签抗体,200 ug 200 ug 235.00 美元 200474 RAT 抗 DYKDDDDK 标签抗体,1 mg 1 mg 399.00 美元 200514 QuikChange 多位点定向诱变试剂盒 30 次反应 1,188.00 美元 200515 QuikChange 多位点定向诱变试剂盒 10 次反应 465.00 美元 200516 QuikChange XL 位点定向诱变试剂盒 30 次反应 855.00 美元 200517 QuikChange XL 位点定向诱变试剂盒 10 次反应 239.00 美元200518 QuikChange 定点诱变试剂盒 30 次反应 608.00 美元 200519 QuikChange 定点诱变试剂盒 10 次反应 239.00 美元 200521 QuikChange II XL 定点诱变试剂盒 10 次反应 239.00 美元 200522 QuikChange II XL 定点诱变试剂盒 30 次反应 608.00 美元 200523 QuikChange II 定点诱变试剂盒 10 次反应 239.00 美元 200524 QuikChange II 定点诱变试剂盒,30 次反应 608.00 美元 200550 GeneMorph II 随机诱变试剂盒 1 套 566.00 美元 200552 GeneMorph II EZClone 域诱变试剂盒 1 套 425.00 美元 200555 QuikChange II-E(电穿孔)定点诱变试剂盒 10 次反应 239.00 美元 200600 DNA 提取试剂盒 1 套 371.00 美元 200820 AccuScript 高保真第一链 cDNA 合成试剂盒 1 套 359.00 美元 201115 Kb DNA Ladder 250 ug 214.00 美元 201190 鲑鱼精子 DNA 1 套 273.00 美元 201222 MiracleHyb高性能杂交溶液,250 mL 250 mL 160.00 $ 204130 GeneJammer 转染试剂,1 mL 1 mL 366.00 $

利用通用供体快速生成可逆和条件等位基因的 HIT 捕获策略
在模型生物中定向诱变是基因功能注释和生物医学研究的关键。尽管 CRISPR-Cas9 系统在基因编辑方面取得了技术进步,但在大型动物模型中快速有效地引入定点突变仍然是一个挑战。在这里,我们开发了一种强大而灵活的插入诱变策略,即同源性独立的靶向捕获 (HIT-trapping),它是通用的,可以有效地靶向捕获内源性目的基因,而不依赖于同源臂和胚胎干细胞。进一步优化并为 HIT-trap 供体配备位点特异性 DNA 倒置机制,可以在单个实验中一步生成可逆和条件等位基因。作为概念验证,我们成功地在原代猪成纤维细胞中为 21 种疾病相关基因创建了突变等位基因,平均敲入频率为 53.2%,比以前的方法有了很大的改进。这里提出的多功能 HIT 捕获策略有望简化突变等位基因的靶向生成,并促进猪等大型哺乳动物的大规模诱变。

新型基因组技术在食品微生物研究中的应用...
欧洲食品和发酵菌种协会 (EFFCA) 和国际益生菌协会 (IPA Europe) 欢迎委员会服务部门为更新欧洲转基因立法而采取的举措。我们同意 2021 年 4 月 29 日发布的欧盟委员会关于新基因组技术 (NGT) 的研究(“根据联盟法律以及欧洲法院在 C-528/16 案中的裁决对新基因组技术的地位进行研究”)的结论,即当前的欧盟转基因框架并不适合目的,没有考虑到新的科学发展和机遇,以解决我们时代的巨大可持续粮食挑战并实现欧盟绿色协议的目标。EFFCA 和 IPA EU 还想回顾一下,新基因组技术开发背后的自然生物学最初是从食品微生物中发现并在工业上加以利用的,正如许多科学出版物所记录的那样(Dal Bello,F. 等人,2024 年)。因此,我们希望呼吁委员会各部门制定一份路线图,对有意发布的立法进行现代化改造,范围包括微生物,包括通过定向诱变和顺式诱变改良的食品培养物。

实时H2O2监视的快速遗传编码传感器的结构引导的工程
A-B结构引导的OROS传感器设计假设。 大肠杆菌的调节结构域(RD)还原和氧化形式的晶体结构。 胱氨酸形金对以黄色标记。 红色指示超级传感器的荧光蛋白插入环,蓝色指示新近鉴定的OROS传感器的荧光蛋白插入位点。 b的氧化氧结构的B因子和残基到残留的距离图,用于放大的假定区域,并在氧化和还原形式的Ecoxyr之间具有高构象变化。 红色和绿色框分别表示HyperRed和Oros-G的插入位点。 针对OROS-G提出的插入位点在C199和C208之间的循环之外(灰色线)。 以最大化循环的灵活性。 OROG-G传感器变体的 C-E筛选。 在HEK293细胞上表达并筛选所有传感器变体(每个条件/变体n> 100个单元)。 c荧光变化(∆F/fo)响应细胞外H 2 O 2(300µm)刺激对CPGFP插入到新鉴定的OROS插入区域的变体上。 插入211-212,确定了具有特殊响应动力学范围的变体。 d插入211-212的最大荧光变化(∆F/fo),并响应高(300µm)和低(10µm)细胞外H 2 O 2。 e位定向诱变变体的最大荧光变化(∆F/fo)预测可减少CPGFP的水获取。 除非另有说明,否则从3个生物学重复中收集利益。A-B结构引导的OROS传感器设计假设。大肠杆菌的调节结构域(RD)还原和氧化形式的晶体结构。胱氨酸形金对以黄色标记。红色指示超级传感器的荧光蛋白插入环,蓝色指示新近鉴定的OROS传感器的荧光蛋白插入位点。b的氧化氧结构的B因子和残基到残留的距离图,用于放大的假定区域,并在氧化和还原形式的Ecoxyr之间具有高构象变化。红色和绿色框分别表示HyperRed和Oros-G的插入位点。针对OROS-G提出的插入位点在C199和C208之间的循环之外(灰色线)。以最大化循环的灵活性。OROG-G传感器变体的 C-E筛选。 在HEK293细胞上表达并筛选所有传感器变体(每个条件/变体n> 100个单元)。 c荧光变化(∆F/fo)响应细胞外H 2 O 2(300µm)刺激对CPGFP插入到新鉴定的OROS插入区域的变体上。 插入211-212,确定了具有特殊响应动力学范围的变体。 d插入211-212的最大荧光变化(∆F/fo),并响应高(300µm)和低(10µm)细胞外H 2 O 2。 e位定向诱变变体的最大荧光变化(∆F/fo)预测可减少CPGFP的水获取。 除非另有说明,否则从3个生物学重复中收集利益。C-E筛选。在HEK293细胞上表达并筛选所有传感器变体(每个条件/变体n> 100个单元)。c荧光变化(∆F/fo)响应细胞外H 2 O 2(300µm)刺激对CPGFP插入到新鉴定的OROS插入区域的变体上。插入211-212,确定了具有特殊响应动力学范围的变体。d插入211-212的最大荧光变化(∆F/fo),并响应高(300µm)和低(10µm)细胞外H 2 O 2。e位定向诱变变体的最大荧光变化(∆F/fo)预测可减少CPGFP的水获取。利益。插入211-212变体的突变E215Y导致了工程OROS-G。描述性统计:误差线和频段代表使用Seaborn(0.11.2)统计绘图套件的中心值趋势的自举置信区间(95%)。

欧洲高级卫生委员会提案...
2023 年 7 月 5 日,欧盟委员会通过了一项提案 2,该提案旨在制定一项关于利用新基因组技术 (NGT) 生产的植物及其食品和饲料产品的新法规,并修订法规 (EU) 2017/625。NGT 是一种新的生物技术,能够对生物体进行有针对性的基因组改变。它们是在 2001 年欧盟现行转基因立法出台后开发的(2001/18/EC 指令)。最著名的 NGT 是 CRISPR-Cas9。欧盟委员会得出结论,现行的转基因立法不再适应这些新发展,不利于开发创新和有益的产品。因此,目前的提案旨在支持欧盟的从农场到餐桌和生物多样性战略。NGT 具有内在潜力,可以通过更快地培育出能够抵御气候变化和瘟疫/疾病的作物来为更可持续的世界做出贡献。欧盟委员会的提案仅涵盖含有来自同一植物(定向诱变)或可杂交植物(同源遗传,包括同源遗传)的遗传物质的植物。转基因植物不包含在内,仍将按照目前的 GMO 法规进行监管。以下两类 NGT 植物被考虑:

利用汇集的 CRISPR 文库对油菜进行基因组规模的靶向诱变
近年来利用CRISPR-Cas9系统构建的二倍体作物突变体文库为功能基因组学和作物育种提供了丰富的资源,然而由于基因组的复杂性,在多倍体植物中实现大规模的定点诱变是一项巨大的挑战。本文证明了利用混合CRISPR文库在异源四倍体油菜中实现基因组规模定点编辑的可行性。共设计了18,414个sgRNA来靶向10,480个目的基因,得到了1104株含有1088个sgRNA的再生转基因植株。编辑询问结果显示,178个基因中93个被鉴定为突变,编辑效率为52.2%。此外,我们发现 Cas9 介导的 DNA 切割倾向于在由同一个 sgRNA 引导的所有靶位点发生,这是多倍体植物中的新发现。最后,我们展示了利用后基因分型植物对各种性状进行反向遗传筛选的强大能力。从正向遗传研究中发现了几个可能主导脂肪酸谱和种子油含量且尚未报道的基因。我们的研究为功能基因组学、优良作物育种提供了宝贵的资源,并为其他多倍体植物的高通量定向诱变提供了良好的参考。

热点-让定向进化更简单 摘要
定向进化已成为一种设计蛋白质各种特性的有力策略。传统的构建文库的方法,如易错的 PCR 和 DNA 改组,通常会产生庞大且效率相对较低的文库。在缺乏高通量筛选方法的情况下,搜索此类文库既费时又费力,而且成本高昂。另一方面,由结构或序列信息引导的定向诱变已成为一种产生所谓智能文库的流行方法。随着有利突变与有害突变比例的增加,智能文库可提高定向进化的效率,前提是靶位预测可靠。突变靶位或热点预测对文库的质量和定向进化的性能至关重要。适当选择热点可以高效合理地生成具有所需特性的蛋白质。本文概述了七种热点,分为两类:基于序列的热点,包括 CbD(保守但不同)位点和共同进化的残基;基于 3D 结构的热点,包括活性位点残基、通道位点、柔性位点、与活性中心偶联的远端位点和界面位点。本综述还介绍了用于识别这些热点的计算工具的最新进展,以及将它们用于酶工程的许多成功案例。