XiaoMi-AI文件搜索系统
World File Search System密码子

问题库 B.Sc III 学期(NEP)
一分 1. 什么是顺反子? 2. 什么是重组? 3. 什么是突变体? 4. 说出 RNA 聚合酶的类型。 5. 说出 RNA 聚合酶的任意两种功能。 6. 什么是转录? 7. 什么是遗传密码? 8. 什么是翻译? 9. 什么是三联体密码子? 10. 什么是起始密码子?举例说明。 11. 什么是终止密码子?举例说明。 12. 什么是摆动假说? 13. 说出翻译所涉及的步骤。 14. 什么是异染色质? 15. 什么是真染色质? 16. 什么是基因表达? 17. 定义封端? 18. 什么是剪接? 19. 定义多聚腺苷酸化。 20. 什么是基因沉默? 21. 什么是 RNAi? 22. 影响基因表达的因素有哪些? (光照、温度、污染) 23. 两种调控基因是什么? 24. 如果基因表达不受调控会发生什么? 25. 什么会刺激基因表达? 26. 什么会增加基因表达? 27. 基因表达的两种主要部分是什么? 28. 基因表达的三个要素是什么? 29. 基因表达的两个阶段是什么? 30. 什么是摆动现象? 31. 什么是糖基化? 32. 什么是甲基化? 33. 什么是磷酸化? 34. 乙酰化是什么意思?

如何治疗KRAS G12C突变的晚期非小细胞肺癌...
在包括 NSCLC 在内的所有肿瘤类型中,约 98% 的致癌 RAS 突变发生在 Switch I 的 G12 或 G13 密码子上,或 Switch II 区域的 Q61 密码子上。24 这些突变的获得导致 KRAS 活性改变,从而维持不受控制的 KRAS 信号网络并促进肿瘤形成和进展(图 1A、B)。KRAS 中的 G12 突变是最常见的突变,占肺癌中所有 KRAS 突变的近 90%,其次是密码子 13 和 61 的突变。24 新兴证据表明,不同的 KRAS 异构体在临床特征、并发基因组变异和基因表达谱方面高度异质,凸显了不同 KRAS 突变体潜在的异构体依赖性治疗脆弱性。 16 KRAS G12C 突变与烟草暴露密切相关,据报道,与其他 KRAS 亚型和 KRAS 野生型 NSCLC 相比,KRAS G12C 突变具有更高的肿瘤突变负担和较高的基因同时突变率,例如 STK11 、 KEAP1 、 SMARCA4 和 ATM 。16,17 此外,具有 KRAS G12C 突变的 NSCLC 倾向于上调免疫逃逸标志物,例如 PD-L1 和 PD-L2,从而部分解释了在该患者群体中观察到的对 ICI 的敏感性增加。16,25
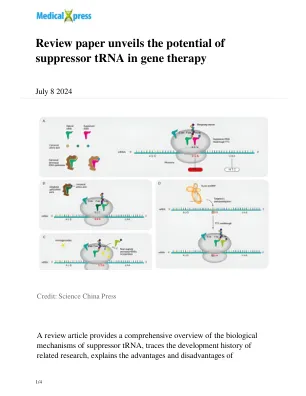
审查论文揭示了基因治疗中抑制器tRNA的潜力
最初,tRNA仅被认为是氨基酸的转运蛋白。通过发现抑制器tRNA发生了变化。1965年,Engelhardt等人。 实验表明,抑制器tRNA可以识别停止密码子和插入氨基酸,从而绕过翻译终止过程。 随后的研究进一步揭示了真核生物中抑制器tRNA的读取机理,并在基因治疗中显示了其潜力。1965年,Engelhardt等人。实验表明,抑制器tRNA可以识别停止密码子和插入氨基酸,从而绕过翻译终止过程。随后的研究进一步揭示了真核生物中抑制器tRNA的读取机理,并在基因治疗中显示了其潜力。

一种新颖的统计方法预测了...的可变性
SARS-CoV-2 病毒已成为 21 世纪最大的流行病,感染人数达数亿,死亡人数达数千万人。世界各地的科学家都在竞相开发疫苗和新药,以战胜这场流行病并为 COVID-19 疾病提供有效的治疗方法。因此,迫切需要更好地了解 SARS-CoV-2 的发病机制如何受到病毒突变的影响,并确定病毒基因组中可作为新疗法稳定靶点的保守片段。在这里,我们介绍了一种文本挖掘方法,可直接从参考(祖先)全基因组序列估计基因组片段的可变性。该方法依赖于根据基因组片段在整个基因组中的空间分布和频率来计算其重要性。为了验证我们的方法,我们对近 80,000 个公开可用的 SARS-CoV-2 前身全基因组序列中的病毒突变进行了大规模分析,并表明这些结果与用于关键字检测的统计方法预测的片段高度相关。重要的是,这些相关性在密码子和基因水平以及基因编码区都成立。使用文本挖掘方法,我们进一步确定了可能成为基于 siRNA 的抗病毒药物候选者的密码子序列。值得注意的是,这项研究中确定的候选者之一对应于刺突糖蛋白表位的前七个密码子,这是唯一一种与人类蛋白质不匹配的 SARS-CoV-2 免疫原性肽。

mRNA-1345的设计和临床前评估...
呼吸道合胞病毒(RSV)是幼儿和老年人下呼吸道疾病的重要原因。我们设计了一种优化的密码子MRNA疫苗mRNA-1345,该密码子编码在预输(PERF)构象中稳定的RSV F-糖蛋白,并在细胞质尾部缺失。mRNA-1345蛋白表达在体外与以前的基于mRNA的RSV疫苗候选物相比临床评估。在啮齿动物模型中,mRNA-1345诱导了强大的中和抗体反应,T辅助1偏置的细胞反应,并证明了针对RSV挑战的剂量依赖性保护,没有呼吸疾病增强的证据。这些发现强调了mRNA-1345作为有效的RSV疫苗的潜力,并通过临床数据证实了mRNA-1345在老年人中的功效。

CRISPR/Cas 与核糖核蛋白复合物和瞬时选择端粒载体可在灰葡萄孢菌中实现高效的无标记和多重基因组编辑
CRISPR/Cas 已成为多种生物体中遗传操作的最先进的技术,能够以前所未有的效率进行有针对性的遗传改变。本文中,我们报告了在重要的坏死性植物病原体灰霉病中首次建立强大的 CRISPR/Cas 编辑,该方法基于将优化的 Cas9-sgRNA 核糖核蛋白复合物 (RNP) 引入原生质体。通过开发一种将 RNP 递送与含端粒的瞬时稳定载体共转化相结合的新策略,进一步提高了编辑产量,从而允许临时选择和方便地筛选无标记编辑事件。我们证明,与现有的基于 CRISPR/Cas 的丝状真菌方法(包括模型植物病原体稻瘟病菌)相比,这种方法提供了更高的编辑率。编辑菌株的基因组测序显示很少有额外的突变,也没有 RNP 介导的脱靶证据。端粒载体介导编辑的高性能通过随机诱变 sdhB 基因的 272 个密码子得到证实,该基因是琥珀酸脱氢酶抑制剂 (SDHI) 杀菌剂抗性的主要决定因素,方法是将 272 个密码子批量替换为编码所有 20 种氨基酸的密码子。在没有选择的情况下,所有交换的频率都相似,但 SDHI 选择允许识别新的氨基酸替换,这些替换赋予了对不同 SDHI 杀菌剂的不同抗性水平。基于 RNP 的共转化效率提高且易于操作,有望加速 B . cinerea 和其他真菌的分子研究。


PiggyBac™转座酶
高活性 piggyBac 转座酶可促进小基因和大基因稳定整合到目标基因组中。该载体不可再生,不能在细菌中繁殖。SPB-100 – 改进的超级 piggyBac 转座酶 mRNA。序列已进行密码子优化(与 SPB-003 相比),以提供更好的 mRNA 稳定性和哺乳动物系统中的蛋白质表达。piggyBac 转座酶 mRNA 适用于无法有效转染载体 DNA 的细胞。SPB-200 – 良好制造级超级 piggyBac 转座酶 mRNA。序列已进行密码子优化,以提供更好的 mRNA 稳定性和哺乳动物系统中的蛋白质表达。piggyBac 转座酶 mRNA 适用于无法有效转染载体 DNA 的细胞。它非常适合将细胞系用于下游制造的应用,即需要 GMP 起始材料的研究和主细胞库。

2025 Ur医学院发现您的SMD周末
Hypermia Susceptibility Authors: Miao He , Sheila Riazi, Luke R. Van Den Berslaar, Gunilla Islander, Luc Heytens, Marc M.J. Snoeck, Andrew Bjorksten, Robyn Gillies, George Drantsar, Anna Hellblom, Susan Treves, Gurrun kunst, Nicol C. Voermans, Heinz Jungbluth, Robert T. Dirksen免疫学,微生物学和病毒学 HIV-1破坏了密码子 - 抗原双工的稳定,以增强核糖体Hypermia Susceptibility Authors: Miao He , Sheila Riazi, Luke R. Van Den Berslaar, Gunilla Islander, Luc Heytens, Marc M.J. Snoeck, Andrew Bjorksten, Robyn Gillies, George Drantsar, Anna Hellblom, Susan Treves, Gurrun kunst, Nicol C. Voermans, Heinz Jungbluth, Robert T. Dirksen免疫学,微生物学和病毒学HIV-1破坏了密码子 - 抗原双工的稳定,以增强核糖体