XiaoMi-AI文件搜索系统
World File Search System对称的

量子热力学的施密特分解方法
摘要:发展量子系统的自洽热力学理论对现代物理学至关重要。尽管它在量子科学和技术中发挥着重要作用,但目前还没有统一的形式来描述一般自治量子系统中的热力学,许多基本问题仍未得到解答。沿着这个思路,大多数当前的努力和方法将分析限制在近似描述和半经典状态的特定场景中。在这里,我们提出了一种基于众所周知的施密特分解来描述任意二分自治量子系统热力学的新方法。这种形式提供了一个简单、精确和对称的框架来表达相互作用系统之间的能量,包括超出标准描述范围的场景,例如强耦合。我们表明,这一过程可以直接识别适合表征物理局部内部能量的局部有效算子。我们还证明这些量自然满足通常的热力学能量可加性概念。

pn7642.pdf
•以高达90 MHz的频率运行•ARM Cortex-M33内置嵌套矢量中断控制器(NVIC)•具有选择源的非掩模中断(NMI)输入,•带有断点和观察点的串行电线调试。包括串行电线输出,以提高调试功能。•系统滴答器•最多21 GPIO(6个专用GPIO)•芯片内存: - 256 kb的闪存(180 kb可供用户使用) - 32 kb RAM(可用于用户使用20 kb)•安全性: - 安全性: - 对称的加密加速器 - 非对称加速器的非对称加速器的密钥 - 固定密钥 - 固定密钥3.用自定义键替换出工厂默认键 - 安全启动支持 - 键传输单元以在密钥存储和加密引擎之间传输对称键,而无需涉及CPU•串行接口 - I 2 C Controller 1

一身纠缠作为费米金系统中的量子资源
我们表明,单身纠缠是纯粹的费米态偏离Slater决定因素(SD)的衡量标准,并由单粒子密度矩阵(SPDM)的混合性确定,可以视为量子资源。相关的理论具有SDS及其凸面作为自由状态,并且保存费米昂线性光学操作(FLO)的数字包括单体统一转换和单粒子模式占用的测量值,作为基本的自由操作。我们首先是基于纯n- fermion态的schmidt样分解的一体纠缠的两拟合公式,可以得出SPDM [与(n-1)体型密度矩阵]从中得出。随后证明,在FLO操作下,初始和计量后的SPDM始终满足主要化关系,从而确保这些操作平均不能增加一体的纠缠。最终表明,该资源与费米子量子计算模型一致,该模型需要超越反对称的相关性。还讨论了更通用的免费测量以及与模式纠缠的关系。

上限在二维电子系统中由微乳液阶段占据的密度窗口
我们提出了一种在不依赖于任何对称性或拓扑的晶格模型中实现零模式的方法,这些对称性或拓扑是对任何类型和强度的大部分中的无序都有坚固的。这种无对称的零模式(SFZM)是通过将带有零模式的单个位点或小群集连接到散装的单个位点或小群集而形成的,该模式用作扩展到整个晶格的“核”。我们确定了该边界与大块之间耦合的要求,这表明这种方法本质上是非遗产的。然后,我们提供了几个示例,这些示例具有任意或结构化的批量,在整体连续体中形成频谱嵌入的零模式,Midgap零模式,甚至还原耦合或障碍转移拓扑拓扑角状态的“ zeroness”。专注于使用光子晶格的可行实现,我们表明,当将光学增益应用于边界时,可以将所得的SFZM视为单个激光模式。
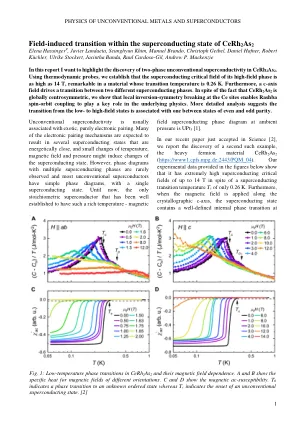
在CERH 2的超导状态中,现场引起的过渡为2 Elena Hassinger#,Javier Landaeta,Seunghyun Khim,Manuel Brando,Christop
Field-induced transition within the superconducting state of CeRh 2 As 2 Elena Hassinger # , Javier Landaeta, Seunghyun Khim, Manuel Brando, Christoph Geibel, Daniel Hafner, Robert Küchler, Ulrike Stockert, Jacintha Banda, Raul Cardoso-Gil, Andrew P. Mackenzie In this report I want to highlight the discovery of two-phase CERH 2 AS 2中的非常规的超导性。使用热力学探针,我们确定其高场相的超导临界场高达14 t,在过渡温度为0.26 k的材料中很有意义。此外,C轴场在两个不同的超导相之间驱动过渡。尽管CERH 2 AS 2是全球中心对称的事实,但我们表明,CE站点的局部反转对称性破坏使Rashba旋转轨道耦合在基础物理学中起关键作用。更详细的分析表明,从低场状态到偶数和奇数之间的过渡与一个状态之间的过渡有关。

生物 Softmax:在现代 Hopfield 网络中得到证明
现代 Hopfield 网络 (HN) 表现出内容可寻址存储器 (CAM) 的特性,可以存储和检索大量记忆。它们还为人类的联想记忆建模提供了基础。然而,这些网络的实现通常在生物学上是不合理的,因为它们假设突触连接的强度是对称的,并使用依赖于多体突触的功能。已经提出了更具生物学现实性的现代 HN 版本,尽管这些实现通常仍使用软最大值函数。计算单个节点的软最大值需要了解所有其他神经元,因此仍然具有一定程度的生物学不合理性。我们提出了一种现代 HN,它使用可以以更符合生物现实的方式计算的 softmax 版本,因此使我们更接近生物学上合理的记忆模型。我们还表明,我们提出的网络可以使用局部学习规则来学习连接权重,该规则源自能量函数的梯度下降。最后,我们验证了我们提出的生物网络的行为类似于现代 HN,并探索了它的其他一些有趣的特性。

超低能耗可逆时间同步量子点细胞自动机组合逻辑电路的设计与仿真
摘要 量子点细胞自动机 (QCA) 代表着一种新兴的纳米技术,有望取代当前的互补金属氧化物半导体数字集成电路技术。QCA 是一种极具前景的无晶体管范式,可以缩小到分子级,从而促进万亿级器件集成和极低的能量耗散。可逆 QCA 电路具有从逻辑级到物理级的可逆性,可以执行计算操作,耗散的能量低于 Landauer 能量极限 (kBTln2)。逻辑门的时间同步是一项必不可少的附加要求,尤其是在涉及复杂电路的情况下,以确保准确的计算结果。本文报告了八个新的逻辑和物理可逆时间同步 QCA 组合逻辑电路的设计和仿真。这里介绍的新电路设计通过使用本质上更对称的电路配置来缓解由逻辑门信息不同步引起的时钟延迟问题。模拟结果证实了所提出的可逆时间同步 QCA 组合逻辑电路的行为,该电路表现出超低能量耗散并同时提供准确的计算结果。

在磁涡场中自旋1/2粒子的超对称性,Anyons
已知具有n = 2超对称性的垂直磁场的量子非偏见自旋1/2平面系统。我们在磁涡流的场中考虑了这样的系统,发现哈密顿量只有两个自我接合延伸与标准n = 2的超对称性兼容。我们表明,只有在这两种情况下,子系统之一与原始的无旋转Aharonov-Bohm模型相吻合,并伴随着超级合作伙伴Hamiltonian,该模型允许波浪函数的单数行为。我们发现了一个额外的非局部运动积分家族,并将它们与局部增压一起在三 - 苏皮对称的统一框架中一起处理。包含动态保形的对称性会导致无限生成的超级级别,其中包含超符号OSP(2 J 2)对称性的几个表示。我们将结果的应用在相同的人的两体模型的框架中。讨论了非平凡的接触相互作用以及新出现的n = 2线性和非线性超对称性。2010 Elsevier Inc.保留所有权利。

学生研究会议
摘要:跨被子植物(开花植物)的证据表明转录因子radialis(rad),divaricata(div)和细胞增多菌(CYC)基因在调节花卉对称性和形态学方面起着重要作用。当前的Snapdragon花卉发育途径模型表明CYC基因控制RAD的激活,从而导致DIV的抑制。跨被子植物(开花植物)的研究表明,双侧对称花的模式受CYC样基因的调节。fedia(caprifoliacea)是双侧对称的花,经过两个重复事件,其中包含CYC2和CYC3的两个副本。我们先前已经表明,这些基因的每个基因,FGCYC2A,FGCYC2B,FGCYC3A和FGCYC3B都会影响该物种中的花卉对称性,并下调CYC样基因表达,从而改变了变为辐射样的花朵。使用这些基因敲除,我们将研究四个CYC旁系同源物对Fedia graciliflora中的Fgrad2和FGDIV1基因的下调。

拉吉夫·甘地 Proudyogiki Vishwavidyalaya, 博帕尔
1. MATLAB 工具介绍。2. 实现连续时间的 delta 函数、单位阶跃函数、斜坡函数和抛物线函数。3. 实现离散时间的 delta 函数、单位阶跃函数、斜坡函数和抛物线函数。4. 实现连续时间的矩形函数、三角函数、sinc 函数和 signum 函数。5. 实现离散时间的矩形函数、三角函数、sinc 函数和 signum 函数。6. 利用代数运算探索信号中偶对称和奇对称的传递。7. 探索信号参数变换(幅度缩放、时间缩放和平移)的效果。8. 探索给定系统的时间方差和时间不变性。9. 探索系统的因果关系和非因果关系。10. 演示两个连续时间信号的卷积。11. 演示两个连续时间信号的相关性。 12. 演示两个离散时间信号的卷积。13. 演示两个离散时间信号的相关性。14. 确定给定信号的傅里叶变换的幅度和相位响应。