XiaoMi-AI文件搜索系统
World File Search System扁虫
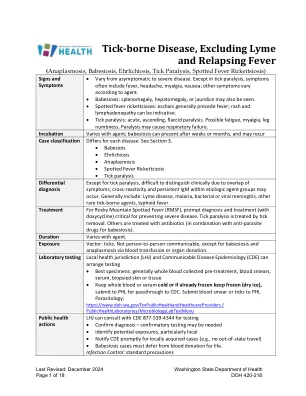
蜱传疾病,不包括莱姆病和回归热
除蜱麻痹外,由于症状重叠,临床上难以区分;可能出现病原体组内的交叉反应和持续性 IgM。通常包括:莱姆病、疟疾、细菌性或病毒性脑膜炎、其他罕见蜱传病原体、伤寒。治疗对于落基山斑疹热 (RMSF),及时诊断和治疗(使用强力霉素)对于预防严重疾病至关重要。蜱麻痹可通过去除蜱虫来治疗。其他可用抗生素治疗(与抗寄生虫药物联合治疗巴贝斯虫病)。持续时间因病原体而异。暴露媒介:蜱虫。除了通过输血或器官捐赠传播的巴贝斯虫病和边虫病外,不会在人与人之间传播。实验室检测当地卫生管辖区 (LHJ) 和传染病流行病学 (CDE) 可以

对种子休眠和发芽中的小麦III类过氧化物酶基因基因锥虫基因的功能分析
怀孕期间的产前超声检查显示出正常的胎儿发育。此外,患者的肾功能正常和血压正常。在妊娠29周时,超声揭示了最深的垂直口袋10厘米的多氢化物。在妊娠33周时,膜和子宫收缩不经常发生过早破裂。然后,进行了紧急的下部CS,并输送了一个重2200克的单个活雌性胎儿。由于呼吸窘迫,该婴儿被送入新生儿重症监护病房(NICU),并接受了补充氧气治疗,最初是通过连续的阳性气道压力(CPAP)持续一天,然后再进行鼻氧。婴儿由专门的肾脏科医生评估,其肾脏和一般检查的结果正常。因此,她出院并跟进,表现出正常的发展里程碑。请求并获得适当的书面知情同意书,并获得了本案报告的个人医学信息。

通过可穿越虫洞协议中的算子扩散实现多体量子隐形传态
通过利用一对量子比特之间的共享纠缠,可以将量子态从一个粒子传送到另一个粒子。最近的进展揭示了量子隐形传态的内在多体泛化,与引力有着巧妙而令人惊讶的联系。具体来说,量子信息的隐形传态依赖于多体动力学,这种动力学源于与引力全息对偶的强相互作用系统;从引力的角度来看,这种量子隐形传态可以理解为通过可穿越虫洞传输信息。在这里,我们提出并分析了一种新的多体量子隐形传态机制——被称为峰值隐形传态。有趣的是,峰值隐形传态利用的量子电路类型与可穿越虫洞隐形传态完全相同,但微观起源却完全不同:它依赖于一般热动力学下的局部算子的扩散,而不是引力物理。我们通过分析和数值方法证明了峰值尺寸隐形传态在各种物理系统中的普遍性,包括随机单元电路、Sachdev-Ye-Kitaev 模型(高温)、一维自旋链和带弦校正的体引力理论。我们的研究结果为使用多体量子隐形传态作为强大的实验工具铺平了道路,用于 (i) 表征强关联系统中算子的尺寸分布和 (ii) 区分一般和内在引力扰乱动力学。为此,我们提供了在捕获离子和里德堡原子阵列中实现多体量子隐形传态的详细实验蓝图;分析了退相干和实验缺陷的影响。

利用 CRISPR/Cas9 系统对原生动物柔嫩艾美耳球虫进行基因改造
摘要 随着反向遗传操作平台的建立,柔嫩艾美耳球虫已成为研究原虫生物学和免疫学的宝贵模式生物。本文介绍了利用CRISPR(成簇的规律间隔的短回文重复序列)/Cas9(内切酶)系统对柔嫩艾美耳球虫进行高效基因编辑的应用,表明CRISPR/Cas9系统可通过一条向导RNA介导位点特异性的双链DNA断裂。利用该系统,我们成功地将红色荧光蛋白插入内源性微线体蛋白2(EtMic2)的C端,对其进行了标记。我们的研究结果将CRISPR/Cas9介导的基因改造系统的应用扩展到柔嫩艾美耳球虫,为针对性地研究顶复门寄生虫的基因功能开辟了一条新途径。

二次胞菌细胞淋巴虫组织细胞增多症治疗转移性子宫内膜癌患者
巨噬细胞激活综合征或血有淋巴细胞培养细胞增多症(HLH)的特征是免疫系统的不适当过度激活,涉及淋巴细胞和巨噬细胞异常的诱导和巨噬细胞的异常(增加)(图。1)[2]。HLH有两种类型:主要和次要。大多数次要形式具有感染性,自身免疫性或医源性病因。诊断需要八个症状中的五个(发烧;脾脏肿瘤;双胆汁症;高甘油三酸酯或低纤维蛋白原血症;胞藻细胞增多症,铁蛋白> 500 µg/l;低/缺乏天然杀手(NK)-cell活性;预后会很快变得严重,死亡率通常很高[4]。不幸的是,临床性质性通常会导致治疗启动的延迟,这是一种治疗性紧急情况,需要病因治疗与免疫靶向治疗相结合以控制发病[5]。此外,它的病理生理学仍然鲜为人知,该综合征的发病率低解释了缺乏标准化治疗[4]。HLH是与ICI治疗相关的罕见不良影响。观测和回顾性研究中报告的发病率<0.1%[6]。尽管在文献中已经描述了与nivolumab,pembrolimab或ipilimumab等ICI相关的几例临床HLH病例,但据我们所知,尚无先前描述过与Dostarlimab治疗相关的临床病例[7]。在这里,我们描述了一名患者,该患者单次注射Dostarlimab后开发了HLH,这是转移性子宫内膜癌的一部分。

综合科学级7周8课2主题
感冒,像艾滋病这样的慢性疾病。除了这些急性疾病外,病毒还负责腮腺炎,脊髓灰质炎,狂犬病等。2。由细菌引起的疾病包括白喉,伤寒,霍乱等。3。疟疾和睡眠疾病是原生动物引起的疾病。4。像round虫一样,tap虫可以分别引起诸如cas虫病和taeniasis之类的疾病。微生物通过以下方式引起疾病:


一开始就进行全编辑
锥虫原生动物参与一些奇怪的生物化学过程,最奇怪的莫过于 RNA 编辑。在这些生物(例如短膜虫、利什曼原虫和锥虫)的线粒体中,蛋白质编码转录本通过位点特异性删除某些基因组编码的尿苷残基并添加其他非编码的 U 而发生改变(参考文献 1)。该过程重新定制初始初级转录本(“预编辑 RNA”),以使最终产品指定完整的功能性蛋白质。除了对预编辑 RNA 的特定区域进行局部编辑(5'-编辑)之外,锥虫还进行一种引人注目的“泛编辑”,在某些情况下,这种编辑可占信使 RNA 成熟序列的 50% 或更多 1 。在本期第 345 页 2 ,Maslov 等人提出的证据表明,与人们的预期相反,泛编辑是锥虫谱系中的古老特性,而非最近获得的特性。RNA编辑系统的进化(它们如何出现以及为何持续存在)是进化生物学中的一个挑战性问题 3 。迄今为止,U添加/删除编辑仅在线粒体中发现,并且仅存在于动基体目(包括锥虫、博多虫和相关的隐虫)中。然而,其他类型的线粒体 mRNA 编辑也已被记录 4 ,以及线粒体转移 RNA S- 7 和核糖体 RNA 8 的编辑。这些系统的多样性和机制独特性,再加上它们的高度受限发生,强烈表明它们中的大多数(如果不是全部)都是最近在进化中获得的特性 3 。
通过胚胎注射和 ReMOT 控制对黑腿蜱(Ixodes scapularis)进行 Cas9 介导的基因编辑
摘要 尽管蜱虫能够获得和传播多种致病病原体,但对蜱虫的研究却落后于蚊子等其他节肢动物媒介,这主要是因为在应用现有的遗传和分子工具方面存在挑战。CRISPR-Cas9 正在改变非模式生物研究;然而,尚未有蜱虫成功进行基因编辑的报道。注射蜱虫胚胎进行基因编辑的技术挑战进一步减缓了研究进展。目前,尚无针对任何螯合动物物种(包括蜱虫)的胚胎注射方案。在此,我们报告了一种针对黑腿蜱(Ixodes scapularis)的成功胚胎注射方案,以及使用此方案通过 CRISPR-Cas9 进行基因组编辑。我们还证明 ReMOT 控制技术可成功用于在昆虫纲之外产生基因组突变。我们的研究结果为蜱研究界提供了创新工具,对于促进我们对蜱虫传播病原体的分子机制以及宿主-媒介-病原体相互作用的潜在生物学的理解至关重要。

比较复杂网格中的Dijkstra和*算法...
●a*算法:在存在燃料站和虫洞时计算出的距离和节点方面表现出了出色的性能。A*的启发式性质使其能够有效利用这些元素,从而导致较短的路径和减少的计算工作。燃料站和虫洞的存在提高了A*的效率,使其可以更快地找到最佳或近乎最佳的路径。●Dijkstra的算法:尽管与**相比,dijkstra的算法通常效率较低,但仍受益于加油站和虫洞。由于燃料站而导致的路径成本的降低以及通过虫洞的可用性提高了其性能,但改进并不像A*那样明显。没有这些元素,Dijkstra的算法在更长的路径和更高的节点计算方面挣扎。