XiaoMi-AI文件搜索系统
World File Search System抗毒
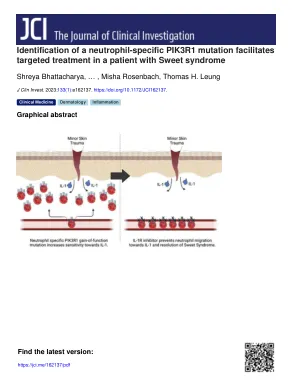
识别中性粒细胞特异性PIK3R1突变促进了甜综合征患者的靶向治疗
患者出现了发烧,神经性,无菌脓尿和右上临界腹部疼痛的其他症状,其疾病炎症都对全身性类固醇治疗做出了反应(图1,B和C)。我们进行了彻底的评估,以排除其表现的其他原因,并评估与甜综合症有关的潜在条件。疾病叶片期间跨激酶和碱性磷酸酶的重复升高导致转介肝病学,肝脏成像证明了原发性硬化性胆管炎的特征,但是血清学检查和流变学评估是自身免疫性和感染的负面性,包括抗病毒性抗肿瘤的抗毒酸及其抗毒胞菌素的负面抗体和替代毒素。肝活检表现出伴有嗜中性炎症的外周纤维化和轻度的胆汁性肠道肠道增殖。对骨髓的检查显示出正常的核型,细胞遗传学和形态。回顾了她先前的肺透明性肉芽瘤病理学和鼻窦活检,排除了IgG4-炎症的疾病。总的来说,我们诊断出患有皮肤和肝介入的Multiorgan甜综合征的患者。

针对 las 或 pqs 群体感应的抗毒药物对囊性纤维化铜绿假单胞菌分离株的体外活性
铜绿假单胞菌引起的慢性肺部感染是囊性纤维化 (CF) 患者发病和死亡的主要原因。针对铜绿假单胞菌群体感应 (QS) 系统的抗毒力药物作为抗生素替代品或佐剂得到了深入研究。之前在非 CF 铜绿假单胞菌参考菌株中进行的研究表明,旧药物氯硝柳胺和氯福克醇可以成功地重新用作分别针对 las 和 pqs QS 系统的抗毒力药物。然而,CF 肺中频繁出现的 QS 缺陷突变体破坏了 QS 抑制剂在 CF 治疗中的应用。在这里,我们在 100 个铜绿假单胞菌 CF 分离株中研究了 QS 信号的产生和对氯硝柳胺和氯福克醇的敏感性,旨在拓宽目前对抗 QS 化合物在 CF 治疗中的潜力的认识。结果表明,我们收集的 CF 分离株中分别有 85%、78% 和 69% 能够熟练使用 pqs、rhl 和 las QS 系统。氯硝柳胺和氯福克醇在体外抑制 QS 和毒力的能力差异很大且因菌株而异。氯硝柳胺的活性范围总体较低,其对 las 信号产生的负面影响与毒力因子产生的减少无关。另一方面,氯福克醇在 CF 分离株中表现出更广泛的 QS 抑制作用,从而降低 pqs 控制的毒力因子绿脓菌素。总体而言,这项研究强调了在进行进一步的临床前研究之前针对大量铜绿假单胞菌 CF 临床分离株测试新型抗毒力药物的重要性,并证实了先前的证据,即 CF 分离株中存在对 QS 抑制剂具有天然耐药性的菌株。然而,研究还表明,对 pqs 抑制剂的耐药性低于对 las 抑制剂的耐药性,从而支持开发 pqs 抑制剂用于 CF 的抗毒力治疗。

五聚体Stx2eB亚单位疫苗诱导猪抗毒抗体及对猪水肿病的预防作用
© 作者 2023。开放存取 本文根据知识共享署名 4.0 国际许可进行授权,允许以任何媒体或格式使用、共享、改编、分发和复制,只要您给予原作者和来源适当的信任,提供知识共享许可的链接,并指明是否做了更改。 本文中的图片或其他第三方资料包含在文章的知识共享许可中,除非资料的致谢中另有说明。 如果资料未包含在文章的知识共享许可中,且您的预期用途不被法定规定允许或超出允许用途,则需要直接从版权所有者处获得许可。 要查看此许可证的副本,请访问 http://creativecommons.org/licenses/by/4.0/ 。知识共享公共领域贡献豁免(http://creativeco mmons.org/publicdomain/zero/1.0/)适用于本文中提供的数据,除非数据来源中另有说明。

Tetanus:您需要知道的
tetanus是一种需要立即治疗的医疗紧急情况,通常包括伤口护理,杀死细菌的药物和抗毒素以中和毒素。任何可能被破伤风细菌污染的伤口的人都应立即寻求医疗救助,尤其是如果他们不确定自己的疫苗接种状况或知道未接种疫苗。即使接受治疗,破伤风也可能是致命的,这就是为什么针对破伤风的疫苗接种非常重要的原因。


在抗高剂量全身性皮质类固醇抗毒表皮坏死式中具有TNF-α抑制剂的阳性经验
史蒂文斯 - 约翰逊综合征(SJS)和有毒表皮坏死溶解(十)是罕见的,潜在的威胁生命的综合症,其特征是坏死性表皮和粘膜病变的发展。SJS/10的最常见病因原因是药物诱导的机制。具有高潜在风险的药物组包括磺酰胺,抗惊厥药,非甾体类抗炎性药物(NSAIDS),别嘌醇,苯巴比妥等。没有针对SJS/10的黄金标准处理算法。在医学实践中,全身性糖皮质激素(SGC),静脉免疫球蛋白(IVIG),血浆置换和环孢菌素在经验上和各种组合中使用。最近发表的研究表明,TNF-α抑制剂是SJS/10的一种有希望的方法,包括对高剂量SGC的抗性病例,依那诺普和二链昔单抗是最常用的药物。在张J等人的大型多中心研究中。 (xxxx),与对照组相比,用依那耐酸,SGCS或两者组合治疗的242例患者的死亡率较低。 与SGC单一疗法相比,皮肤愈合时间较短,从而降低了继发感染的风险。 已发布的数据显示,THF-α抑制剂阻滞具有很高的效率,但是由于缺乏可用信息,因此SJS/TEN患者中TNF-α抑制剂的安全性仍然值得怀疑。 由于应积累所有临床研究数据,以提供可靠的证据,即SJS/TEN的使用TNF-α抑制剂可能是有益的,因此我们报告了一个与Etoricoxib相关的SJS的案例,在50岁的女性中,该病例与十岁的女性相关,而这是50岁的女性,该女性是饮食来饮食高剂量SGCS治疗的案例。在张J等人的大型多中心研究中。(xxxx),与对照组相比,用依那耐酸,SGCS或两者组合治疗的242例患者的死亡率较低。与SGC单一疗法相比,皮肤愈合时间较短,从而降低了继发感染的风险。已发布的数据显示,THF-α抑制剂阻滞具有很高的效率,但是由于缺乏可用信息,因此SJS/TEN患者中TNF-α抑制剂的安全性仍然值得怀疑。由于应积累所有临床研究数据,以提供可靠的证据,即SJS/TEN的使用TNF-α抑制剂可能是有益的,因此我们报告了一个与Etoricoxib相关的SJS的案例,在50岁的女性中,该病例与十岁的女性相关,而这是50岁的女性,该女性是饮食来饮食高剂量SGCS治疗的案例。


使用果皮对肉鸡生长和健康的影响
为了确保肉鸡养殖的长期生存能力,生产商必须解决许多问题,包括饲料成本上升,禁止抗生素增长促进剂以及消费者对肉鸡肉肉化学残留物的认识日益提高。果皮是一种没有商业价值的废物,但是由于其较高的营养含量,尤其是在能源方面,它有可能用作肉鸡小鸡的替代饲料来源。果皮还含有许多营养化合物,由于其抗菌,抗氧化剂和免疫刺激性特性,有可能添加饲料或用作肉鸡的天然补充剂。果皮具有较高的纤维含量以及抗毒和有毒成分,可能会干扰肉鸡的消化和生理功能,因此应谨慎使用肉鸡生产。各种过程,包括发酵,提取,蒸馏以及与其他活性成分(例如酶)结合,可用于优化在肉鸡生产中使用果皮的使用。本评论研究了果皮的使用及其对肉鸡生长和健康的影响。

母婴之家调查委员会-审判...
接种疫苗后的随访 接种疫苗后,每两天对所有儿童进行一次观察,持续两周。接种疫苗九周后,对另外 46 名儿童进行了 Schick 测试。Schick 测试是一种特定的皮肤测试,用于确定一个人是否易患白喉。将少量(0.1 毫升)稀释的白喉毒素注射到皮下以引起反应。如果没有反应,则表明具有免疫力(免疫系统将能够抵御疾病)。在接受 Schick 测试的 46 名儿童中,45 名没有反应,一名儿童有反应。接种疫苗后,除了所有儿童都接受的 Schick 测试外,还随机选择了 12 名儿童进行血液测试。Schick 测试结果呈阳性的一名儿童血液中的抗毒单位水平也很低,这表明他们对疫苗的反应不佳。

目前的肝纤维化治疗研究
长期肝损伤导致肝纤维化,经常发展为肝硬化,肝衰竭,门静脉高血压和肝细胞癌。目前尚无有效的疗法可用于肝纤维化。因此,正在进行抗毒疗法的持续研究。近年来,抗蛋白研究的主题是根据目前对疾病病理的理解,基于原理的治疗分子选择。研究工作主要是为了重新利用目前的FDA批准的药物,旨在涉及肝脏纤维化涉及的病因学分子因素。同时,研究还集中在实验性的小分子上,并有证据表明或扭转纤维化。天然化合物,免疫学和遗传方法已显示出显着的令人鼓舞的作用。本综述总结了针对各种分子靶标的当前抗纤维化药物的效率和安全性,以及抗纤维化药物的特性,主要是在II期和III期临床试验中。