XiaoMi-AI文件搜索系统
World File Search System折叠的

基于纳米的AD和PD
最普遍的神经退行性疾病是阿尔茨海默氏病(AD)和帕金森氏病(PD)。AD和PD均被归类为蛋白质病,在PD中发现了AD中错误折叠的淀粉样蛋白β和α-突触核蛋白中的tau蛋白。主要的广告标志是记忆丧失,在PD中发现多巴胺能神经元的丧失和Lewy-Bodies的发展。运动神经元活性中的缺陷在多巴胺能神经元失去50-70%后,可以注意到NABESIA NIGRA(SN)地区。新兴证据表明,错误折叠的蛋白质缠结和/或斑块具有prion样蛋白,这是引起发病机理的主要因素。可能影响这些疾病病理的其他因素包括氧化应激,线粒体损伤,炎症和与年龄相关的细胞死亡。慢性炎症也被普遍认为在PD的启动和发展中起着核心作用。

使用高阶的简单复杂传染
摘要 - 神经退行性疾病的特征是复杂的蛋白质错误折叠的大脑内传播。例如,当前的发现突出了2种特定错误折叠蛋白在阿尔茨海默氏症中的作用,这些蛋白质被认为使用脑纤维作为高速公路传播。先前的研究通过模拟模型或基于机器学习的预测变量调查了这种扩散,这些预测因素采用大脑连接组作为基础扩散网络。但是,构造的结构连接组仅描述图中节点之间的成对连接。高阶相互作用复杂网络比正常图提供了显着的优势,因为它们可以捕获超出简单的成对关系船的交互。蛋白质错误折叠和聚集通常涉及正常图的合作行为或群体动力学,其专注于单个边缘,无法充分代表。蛋白质错误折叠的非线性和多尺度可能更适合更丰富的高阶模型。在这项研究中,我们研究了高阶网络在这种情况下是否可以提供改进的拟合和解释能力。更具体地说,我们采用淀粉样蛋白β的简单复杂传染模型来预测蛋白质错误折叠的扩散。Simplicial Cronagion复合物在2年的地平线和其他结果中,阿尔茨海默氏症患者在所有大脑区域的预测蛋白质沉积中产生了0.030的平均重建误差,胜过先前的研究,尤其是对于错误折叠的蛋白质的病例稳定增长。尽管时间范围有限,但这项研究突出了结合先进网络分析以捕获跨神经网络蛋白质聚集的复杂动力学的潜力。临床相关性 - 这项研究突出了高阶网络在阿尔茨海默氏症中提高错误折叠蛋白传播的预测的潜力,从而更好地洞悉了蛋白质聚集动力学。

帕金森病治疗的新进展
• 被动提供针对 α -突触核蛋白的抗体 • 让身体产生自己的 α -突触核蛋白抗体(即 α -突触核蛋白疫苗) • 用小分子干扰 α -突触核蛋白 • 创造一种破坏错误折叠的 α -突触核蛋白的蛋白质

非线性物理系统中隐藏的多稳定性
在非线性物理系统中识别逃避直接实验检测的隐藏状态很重要,因为干扰和噪音可以将系统置于隐藏状态,并带来有害后果。我们研究了一个空腔岩石系统,其主要物理学是光子和镁kerr效应。在数值实验中扫描分叉参数(如在实际实验中所做的那样)导致具有两个不同稳定稳态状态的磁滞回路,但是分析计算在环路中赋予了第三个折叠的稳态“隐藏”,这导致了隐藏可粘性的现象。我们提出了一种实验可行的控制方法,将系统驱动到折叠的隐藏状态中。我们通过三元腔镁质系统和基因调节网络证明了这种隐藏的多稳定性实际上很普遍。我们的发现阐明了非线性物理系统中隐藏的动力状态,这些状态不是直接观察到的,但可以在应用中带来挑战和机遇。

国际研究出版与评论杂志-IJRPR
全球数百万人患有神经退行性疾病(NDDS),这是一组神经系统疾病,其标志着重要的中枢神经系统(CNS)或周围神经系统(PNS)中神经元的逐渐丧失。由于神经元网络被终止区分,因此由于神经元死亡,神经网络失去结构和功能,它们无法成功恢复自己。这种干扰会影响基本的沟通途径,这无疑会导致行为,记忆,认知,感觉知觉和/或运动技能的问题。蛋白质折叠和错误折叠的关键过程确立了蛋白质在细胞内的作用或位置。要使许多活性蛋白正确起作用,它们必须形成分组或低聚物。由小管蛋白和肌动蛋白等结构蛋白组成的复杂系统对于多种细胞功能至关重要。许多细胞过程取决于这些分组和系统的高度调节。但是,错误折叠的蛋白质会产生危险的,不受控制的簇,引起许多疾病。蛋白质错误折叠式疾病(PMD),包括阿尔茨海默氏病(AD),帕金森氏病(PD)和病毒疾病,是由通常在体内正确折叠的蛋白质引起的与年龄相关的疾病。众所周知,核酸(NAS)可能与容易发生淀粉样蛋白相互作用以促进聚集过程。多种化学接触,包括由水分子介导的氢键,非极性相互作用和疏水力,参与蛋白质与NAS之间的相互作用。

人类大脑类器官的力学
人类大脑特有的回旋形状最早出现在埃德温·史密斯纸莎草书中,这是一份可追溯到公元前 1700 年的埃及手稿,其中将大脑回旋与熔融金属中的波纹或皱纹进行了比较 [1]。自 19 世纪初以来,这些回旋的描述、发展和功能也一直是研究的主要课题 [2]。回旋的可见上部称为脑回,其深沟称为脑沟。从几何学上讲,回旋增加了给定体积的大脑的表面积。从功能角度来看,它们被认为具有增加皮质内神经元体数量和促进神经元之间连接从而减少电信号在不同区域之间传输时间的战略功能。尽管人们提出了不同的解释,但脑回形成背后的机制尚未完全了解。现在人们普遍认为,人类大脑折叠的出现是内在的机械力而不是外部约束[3]。最近的观察性研究[4,5]进一步支持了皮质在发育过程中快速切向扩张是折叠的主要驱动力[2,6-9]。 44 从最简单的物理层面上讲,折叠的开始可以理解为压缩的上皮层中弹性能量的初始积累,以及薄膜和基底的褶皱变形部分释放。实验中,这种不稳定性可以在与弹性盘结合的圆形壳的受限聚合物膨胀中观察到,这引发了相同类型的褶皱模式[10-14]。在由具有不同膨胀特性的聚合物凝胶制成的双层大脑原型52上进行的类似实验53再现了与真实大脑的脑回和脑沟相似的褶皱54[15]。55

低氧诱导因子2α在肺癌中的作用(综述)
摘要。内质网(ER)是用于蛋白质合成,折叠和修饰,脂质合成和钙储存的必不可少的细胞器。当内源性或外源性刺激导致ER合成的蛋白质折叠功能障碍时,许多展开或错误折叠的蛋白会积聚在ER腔中,并引起一系列随后的反应,称为ER应激。如果ER应力是连续的,则展开的蛋白质反应(UPR)不足以去除累积的展开和错误折叠的蛋白质,因此,UPR信号通路将驱动细胞凋亡。胶质母细胞瘤(GBM)目前是神经系统中最具侵略性和最常见的恶性肿瘤。由于ER应力可能会增加GBM对替莫唑胺的敏感性,因此本文回顾了ER应激诱导的凋亡的可能机制和影响ER应激的因素,并评估了ER应激作为治疗靶标的潜力。

肝细胞癌的耐药性和内质网应激
摘要:肝细胞癌 (HCC) 是全球最常见和最致命的癌症之一。它通常在晚期才被诊断出来,其特点是具有较高的内在耐药性,导致化疗效果有限和治疗后复发。因此,迫切需要了解导致耐药性的潜在机制,并制定克服这一问题的治疗策略。肿瘤细胞的快速增殖,加上高度炎症的微环境,导致不同肝细胞群中蛋白质合成的慢性增加。这导致对蛋白质折叠的需求增加,这不可避免地导致错误折叠或未折叠的蛋白质在内质网 (ER) 腔内积聚。这个过程称为 ER 应激,它会触发未折叠蛋白反应 (UPR) 以恢复蛋白质合成,或者在严重或长期 ER 应激的情况下诱导细胞死亡。有趣的是,ER 应激信号通路的三个不同分支已被证实能够驱动多种肿瘤产生化学耐药性,因此可能成为有希望的治疗靶点。本综述概述了 ER 应激和 UPR 激活如何导致 HCC 产生耐药性。

AffLecto:一个生成Alphafold模型柔性蛋白质构象合素的Web服务器
本质上无序的蛋白质和区域(IDP/IDR)利用其结构性挠性来实现必要的细胞功能,并且功能障碍通常与严重疾病有关。然而,它们的序列,结构动力学和功能角色之间的关系仍然很少理解。将这些复杂关系熟悉的对于特性的发展至关重要,强调了对产生质量IDP/IDR构象异构体的方法的需求。 虽然Alphafold(AF)在建模结构域中表现出色,但它无法准确地代表无序区域,而蛋白质组织的很大一部分不准确。 我们提出了Afflecto,这是一种用户友好的Web服务器,用于生成蛋白质的大构象合奏,其中包括AF结构模型的结构化域和IDR。 通过分析其结构上下文,将IDR识别为尾部,接头或循环。 此外,它结合了一种方法来识别有条件折叠的IDR,AF可能错误地预测为本质上折叠的元素。 使用有效的随机采样算法在全球探索构象空间。 AffLecto的Web界面允许用户通过修改有序区域和分配区域之间的边界以及在几种采样策略之间进行选择来自定义建模。 Web服务器可在https://moma.laas.fr/applications/afflecto/免费获得。对于特性的发展至关重要,强调了对产生质量IDP/IDR构象异构体的方法的需求。虽然Alphafold(AF)在建模结构域中表现出色,但它无法准确地代表无序区域,而蛋白质组织的很大一部分不准确。我们提出了Afflecto,这是一种用户友好的Web服务器,用于生成蛋白质的大构象合奏,其中包括AF结构模型的结构化域和IDR。通过分析其结构上下文,将IDR识别为尾部,接头或循环。此外,它结合了一种方法来识别有条件折叠的IDR,AF可能错误地预测为本质上折叠的元素。使用有效的随机采样算法在全球探索构象空间。AffLecto的Web界面允许用户通过修改有序区域和分配区域之间的边界以及在几种采样策略之间进行选择来自定义建模。Web服务器可在https://moma.laas.fr/applications/afflecto/免费获得。
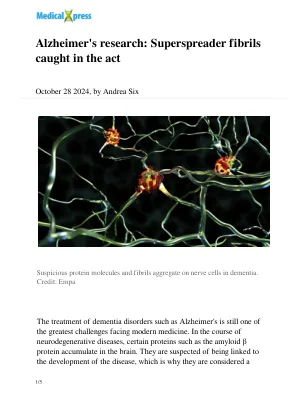
阿尔茨海默氏症的研究:被捕获的超级主架
已知错误折叠的淀粉样蛋白β蛋白的化学组成。以前尚不清楚蛋白质构建块如何组合在一起形成第二代纤维及其形状和结构的机制。“常规方法,例如基于染色技术的方法,可以改变蛋白质的形态和吸附位点,以便无法以自然形式进行分析。” Nirmalraj说。