XiaoMi-AI文件搜索系统
World File Search System斑马

斑馬魚@巴斯
斑马鱼@巴斯 您是否和我们一样对斑马鱼研究充满热情?您想在联合国教科文组织世界遗产城市生活和工作吗? https://whc.unesco.org/en/list/428/ 那就来巴斯大学吧,巴斯大学是一所全球排名前 150 的大学(QS 2025) https://www.topuniversities.com/qs-top-uni-wur 我们的研究人员 Philip Ingham 教授 FRS Philip 在英国率先使用斑马鱼作为模型生物,早在 1980 年代就在牛津大学建立了第一个斑马鱼研究实验室。从那时起,他在 CRUK 伦敦研究所、谢菲尔德大学和埃克塞特大学以及新加坡李光前医学院建立了设施。他曾担任国际斑马鱼学会主席和斑马鱼疾病模型学会副主席,在 Hedgehog 信号通路和斑马鱼骨骼肌发育方面做出了重要发现。他于 2005 年荣获遗传学会奖章,并于 2014 年荣获 BSDB 沃丁顿奖章。罗伯特·凯尔什教授罗伯特在剑桥大学学习进化发育生物学,后与图宾根马克斯物理研究所的 Christiane Nüsslein-Volhard 和俄勒冈大学的 Judith Eisen 一起从事斑马鱼博士后研究。他的研究重点是神经嵴细胞的发育,特别是命运决定。他采用了从 CRSPR-Cas9 介导的基因组编辑到数学建模等一系列方法来剖析转录因子及其相关基因调控网络在选择和平衡命运决定中的作用。去年,他的研究成果获得了国际色素细胞学会联合会 (IFPCS) 的 2023 年迈伦·戈登奖巴斯全球讲席教授 Steven Farber Steve 是约翰霍普金斯大学脂质代谢和功能领域的世界知名专家,他因客座教授的身份定期来巴斯访问。获得电气工程学位后,Steve 在麻省理工学院学习神经生物学,探索胆碱能脑区神经递质和膜磷脂合成之间的平衡。在卡内基研究所 Marnie Halpern 实验室从事博士后研究期间,他率先使用斑马鱼进行脂质生物学研究。他研究的一个主要主题是开发工具,以研究完整组织和器官中脂质的细胞生物学,而这种方式以前只能在培养细胞或酵母中实现。副教授 Vasanta Subramanian 以研究哺乳动物发育而闻名,她从哥廷根 MPI Peter Gruss 实验室的研究员开始研究哺乳动物发育,Vasanta 拥有更多


Archimer-Ifremer公司
转基因斑马鱼模型可有效用于研究内分泌干扰化学物质 (EDC) 的影响;从而了解其作用机制。然而,鉴于已报道的斑马鱼品系在遗传、生理和行为层面存在差异;在使用这些转基因模型进行 EDC 测试之前应谨慎行事。在本研究中,我们在不同的转基因和/或突变斑马鱼品系中进行了一系列实验,以进行 EDC 测试:casper、cyp19a1a-eGFP、cyp19a1a-eGFP-casper、cyp11c1-eGFP、cyp11c1-eGFP-casper。评估了一些常用于 EDC 测试的行为特征以及一些生化和生殖生理终点,并将其与 WT AB 斑马鱼中获得的进行比较,以确保转基因插入和/或突变不会对鱼的基础生殖生理或行为产生负面影响。
使用安全评估向导打印Android的DNA打印机设置实用程序
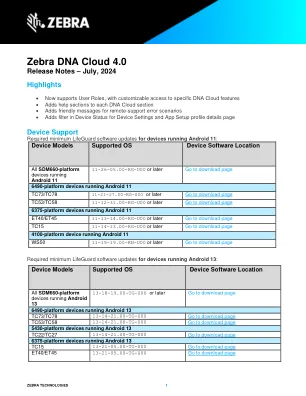
斑马DNA云将我们的迁移率DNA投资组合解决方案集成到一个直观组织的界面中。通过利用我们现有软件的功能,我们为客户和合作伙伴提供了帮助部署,管理和支持设备生命周期的每个阶段的解决方案。使用斑马DNA云,组织可以通过利用移动性扩展(MX)来量身定制设备设置以满足其特定需求。应用程序管理和移动性DNA工具的配置将确保用户可以访问完成其工作所需的斑马应用程序。管理员可以利用救生员OTA来确保其设备始终具有最新功能和安全性,所有这些都单击,而无需物理触摸每个设备。

识别重复的SOX因子的功能分子基础,控制斑马鱼中的内胚层形成和左右模式
确定重复的SOX因子功能差异的分子基础,控制斑马鱼Simaran Johal 1,Randa Elsayed 2,Kristen A. Panfilio 1,3,4,Andrew C. Nelson 1* 1* 1.生命科学学院,吉布特山校园,沃里克大学,考文垂,Cv4 7al,英国2。Warwick医学院,Gibbet Hill校园,沃里克大学,考文垂,Cv4 7al,英国3。 霍恩海姆大学生物学研究所分子遗传学系。 30,70599德国斯图加特4。 动物学研究所:发育生物学,科隆大学,苏黎世大学,苏黎世大学47b,50674德国科隆 *通讯作者电子邮件:a.nelson.1@warwick.ac.ac.ac.ac.ac.ac.ac.ac.ac(acn)摘要内科胰腺。 这些器官的放置和图案依赖于左右的组织者 - 斑马鱼中称为库普弗囊泡(KV)。 转录因子Sox32和Sox17是斑马鱼Soxf亚科的成员。 Sox32和Sox17来自Teleost血统中祖先Sox17的重复。 SOX32在早期胚胎中诱导SOX17的表达,是内胚层和KV祖细胞规范所必需的。 斑马鱼Sox17与KV形态发生有关。 在哺乳动物中,Sox17对于内胚层形成至关重要,可以诱导内胚层祖细胞身份。 因此,表型证据表明斑马鱼Sox32和Sox17与哺乳动物Sox17之间的功能相似性。Warwick医学院,Gibbet Hill校园,沃里克大学,考文垂,Cv4 7al,英国3。霍恩海姆大学生物学研究所分子遗传学系。30,70599德国斯图加特4。动物学研究所:发育生物学,科隆大学,苏黎世大学,苏黎世大学47b,50674德国科隆 *通讯作者电子邮件:a.nelson.1@warwick.ac.ac.ac.ac.ac.ac.ac.ac.ac(acn)摘要内科胰腺。这些器官的放置和图案依赖于左右的组织者 - 斑马鱼中称为库普弗囊泡(KV)。转录因子Sox32和Sox17是斑马鱼Soxf亚科的成员。Sox32和Sox17来自Teleost血统中祖先Sox17的重复。SOX32在早期胚胎中诱导SOX17的表达,是内胚层和KV祖细胞规范所必需的。斑马鱼Sox17与KV形态发生有关。在哺乳动物中,Sox17对于内胚层形成至关重要,可以诱导内胚层祖细胞身份。表型证据表明斑马鱼Sox32和Sox17与哺乳动物Sox17之间的功能相似性。我们试图探索斑马鱼早期胚胎中这些蛋白质之间的功能差异和潜在的相似性。我们的结果表明,与Sox32不同,人类Sox17不能在斑马鱼中诱导内胚层规范。此外,使用混合蛋白功能分析,我们表明SOX32内胚层基因调节网络的特异性与其HMG域与旁产皮sox17的进化差异有关。此外,Sox32和Sox17的C末端区域的变化是其在介导差异基因调节程序中的不同目标特异性和差异的基础。最后,我们确定C末端结构域中的特定保守肽对于SOX17在建立正确的器官不对称性中的作用至关重要。总体而言,我们的结果为脊椎动物内胚层发育,左右模式以及SOXF转录因子功能的演变提供了新的见解。

翻译 GWAS 识别的心律基因座和……
一项全基因组关联研究 (GWAS) 的荟萃分析确定了八个与心率变异性 (HRV) 相关的基因座,但这些基因座中的候选基因仍未得到表征。我们开发了基于图像和 CRISPR/Cas9 的流程,系统地表征活斑马鱼胚胎中 HRV 的候选基因。在转基因表达平滑肌细胞 GFP 的斑马鱼 (Tg[ acta2:GFP ]) 的卵子中同时靶向六个人类候选基因的九个斑马鱼直系同源物,以使跳动的心脏可视化。在受精后 2 天和 5 天,对 381 个活的完整斑马鱼胚胎中的心房跳动进行 30 秒重复记录的自动分析突出显示了影响 HRV 的基因( hcn4 和 si:dkey-65j6.2 [KIAA1755] );心率( rgs6 和 hcn4 );以及窦房停顿和骤停风险( hcn4 )。暴露于 10 或 25 µM 伊伐布雷定(HCN 的开放通道阻断剂)24 小时后,在受精后 5 天,剂量依赖性地导致 HRV 升高和心率降低。因此,我们的筛选证实了已确定的心率和节律基因(RGS6 和 HCN4)的作用;表明伊伐布雷定可以降低斑马鱼胚胎的心率并增加 HRV,就像在人类中一样;并突出了一个在 HRV 中发挥作用的新基因(KIAA1755)。

围产期产妇应力对孕产妇和婴儿肠道和人乳微生物的影响:范围审查方案
斑马鱼是被选为实验室使用的流行研究生物,部分原因是宠物贸易的广泛差异。许多实验室斑马鱼的当代菌落都保存在水产养殖设施中,这些设施监测并旨在遏制可能会对菌落健康和混淆实验产生负面影响的感染。与宠物贸易相比,在研究环境中,实验室控制对与斑马鱼相关的微生物成分的影响尚不清楚。在两个环境中,未知原因的疾病都是常见的。我们进行了一项元文字组合调查,以广泛比较宠物贸易和实验室环境中斑马鱼相关的微生物。我们从宠物贸易中发现了许多微生物在实验动物中未发现的微生物。同居实验揭示了几个可传播的微生物,包括新描述的未发育的,duble stanand的RNA病毒,在Birnaviridae家族中,我们将Rocky Mountain Birnavirus(RMBV)命名。感染,但是当传播到实验动物时,rmBV与明显的抗病毒药反应和出血性疾病有关。这些实验将宠物贸易作为与斑马鱼相关的不同微生物的独特来源,并为发现新描述的病原病毒和其他传染性微生物的发现范式建立了范式,这些微生物可以在实验室中进行研究。

TC5X配置和配件指南
TC5X触发器手柄附件为用户提供了明显的灵活性,因为他们将TC5X设备调整到零售环境中的不同操作中。TC5X触发器手柄附件被认为是一种易于使用的物品,在TC5X设备的使用寿命中可能需要更换。因此,斑马强烈建议购买斑马触发器手柄服务计划。

聚苯乙烯微塑料对斑马鱼胚胎学的影响:两种不同尺寸的比较
微塑料已成为全球一个巨大的问题,因此,研究其对人类和环境健康的可能影响很重要。在这项研究中,斑马鱼胚胎分别比较了两种不同尺寸的聚苯乙烯微塑料(PS -MP),分别为1 µm和3 µm,在0.01、0.1、1.0、1.0和10.0 mgl -1时,并监测高达72小时。毒性测试表明,PS-MP都没有改变胚胎的存活率和正常的孵化过程。相反,两种大小的浓度较高,导致心率和表型变化的增加。以10.0 mgl -1的浓度在幼虫中输入和积累的两个大小的PS -MP,相同的浓度导致凋亡过程的增加与氧化还原稳态变化相关。报告的结果对暴露于PS-MP的负面影响并提供了有关其毒性的新信息的现实看法,也考虑了其尺寸。

微生物从宠物店到实验室繁殖的斑马鱼的传播揭示了一种致病性Birnavirus
新的免疫检查点正在出现,以提高对免疫药物的反应率。由于参与肿瘤微环境的免疫抑制,腺苷A 2A受体(A 2A R)被提议作为免疫发育的靶标。封锁2A R可以恢复肿瘤免疫力,从而改善患者的预后。在这里,我们描述了通过噬菌体显示的人A 2A R(HA 2A R)的有效,选择性和抑制肿瘤抗体拮抗剂的发现。We con- structed and screened four single-chain variable fragment (scFv) libraries—two synthetic and two immunized—against hA 2A R and antagonist-stabilized hA 2A R. After biopanning and ELISA screening, scFv hits were reformatted to human IgG and triaged in a series of cellular binding and functional assays to identify a lead candidate.铅候选者TB206-001散布了HA 2A R-Over表达HEK293细胞的纳摩尔结合;与小鼠和cynomolgus a 2a r的交叉反应性,但不是人类A 1,2b或3受体; HA 2A R在HA 2A R-r-evercress表达HEK293细胞和外周血单核细胞(PBMC)中的功能拮抗作用;结肠肿瘤的HUCD34-NCG小鼠中的肿瘤抑制活性。鉴于其治疗特性,TB206-001是将其纳入下一代双特异性免疫治疗药的良好候选者。