XiaoMi-AI文件搜索系统
World File Search System末端的

末端舒张和终端施加压力的物理模型 -
左心室刚度和收缩力,其特征是末期压力 - 卷卷关系(EDPVR)和末端的2骨压力 - 卷量关系(ESPVR),是人心脏表现的两个重要指标。尽管已经对EDPVR和ESPVR进行了大量研究,但是没有介绍具有结合两种关系的物理解释参数的模型,从而损害了对心脏生理和病理学的理解。在这里,我们提出了一个模型,该模型在统一框架中对参数进行物理解释评估EDPVR和ESPVR。我们的基于物理的模型拟合了可用的实验数据,并且在计算机结果中非常符合现有模型的表现。带有规定的参数,新模型用于预测左心室的压力量关系。我们的模型对心脏力学有了更深入的了解,因此将在心脏研究和临床医学中应用。

端粒酶作为...
端粒是每个染色体末端的重复,非编码DNA序列(Louzon等,2019)。端粒酶是一种核糖核蛋白酶,活性为80% - 90%的恶性肿瘤,其功能是维持染色体端粒(Arndt和Mackenzie,2016年)。端粒酶复合物由两个成分组成,即催化亚单位人类端粒酶逆转录酶(HTERT)和端粒RNA成分(HTERC)(Peska and Garcia,2020年)。端粒酶在大多数正常的人类细胞中都是由于严格的转录抑制而无效的。然而,它的激活被认为是人类细胞恶性转化的先前步骤(两者,2017年)。此外,HTERT除了其在端粒延长方面的功能外,还具有其他明显的生物学活性。例如,它保护癌细胞免受化学治疗药物诱导的凋亡。此外,高端粒酶表达使癌细胞抗化疗和

Neuropilin-1抑制作用抑制神经生长因子...
引言神经生长因子(NGF)的刺激能够增强交感神经元的生长的能力(1)。tovyosin相关激酶A(TRKA),一种受体酪氨酸激酶(RTK),介导了NGF的神经营养作用(2)。在NGF与周围神经末端的TRKA结合后,NGF/TRKA信号体被恢复到SOMA,在那里它们调节了转换(3,4)。p75 NTR,NGF和Pro-NGF的受体(5,6),作用促凋亡信号通路(7)。NGF和TRKA也介导疼痛(8)。尽管在神经元发育和疼痛的背景下对NGF及其受体进行了深入的研究,但对NGF信号传导的理解不足阻碍了对NGF指导的治疗剂的认可。慢性疼痛遭受了百分之二十的人口,但受到非甾体类抗炎药和阿片类药物的治疗不足,这些药物缺乏疗效并具有危及生命的副作用。
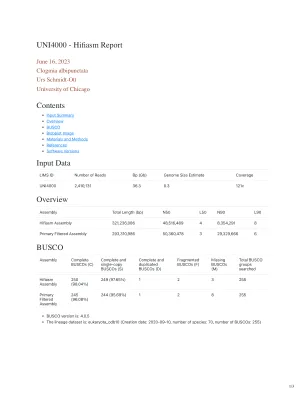
输入数据概述BUSCO UNI4000 -CDN
对于每个DiDail omni-c文库,将染色质与甲醛固定在原子核中,然后提取。用DNase I消化了固定的染色质,将染色质末端修复并连接到生物素化桥适配器,然后将含有末端的衔接子接近粘合。接近连接后,将交联后逆转并纯化了DNA。纯化的DNA以去除未结扎片段内部的生物素。使用NEBNEXT Ultra酶和Illumina兼容适配器生成测序文库。在每个文库富集之前,使用链霉亲和素珠分离含生物素的片段。库是在Illumina Hiseqx平台上测序的,以产生约30倍的序列覆盖率。然后Hirise使用MQ> 50读脚手架的读数(有关数字,请参见上面的“读取对”)。

研究生生物化学/生物化学1发现...
在过去的二十年中,基于荧光蛋白的生物传感器已成为使用显微镜使用空间和时间分辨率的细胞种群监测和测量细胞氯化物的重要工具。由于氯化物转运蛋白在各种生物学过程中的无处不在,研究人员必须配备具有一系列特性的生物传感器。这些包括激发/排放曲线,氯化物结合亲和力和操作pH值。然而,该末端的进展仅限于单个域,强度计和多域,比率传感器。为了扩展氯化物生物传感器工具包,我们正在识别和设计用于细胞成像应用的新的氯化物敏感荧光蛋白。在本演讲中,我将描述1)结构引导的生物信息学如何导致从Hydrozoa clytia glytia Clytia Clytia和2)指导进化如何解锁单个领域,激发率ratiememetric传感器。

生成Zeb2不足的人IPSC线(...
智力残疾,癫痫,赫希斯普朗氏病和各种先天性畸形(Garavelli and Mainardi,2007年)。此外,Zeb2的过表达与不同形式的癌症的进展有关(Fardi等,2019)。虽然已经对Zeb2蛋白的功能进行了广泛的研究,但目前缺乏可用的Zeb2缺乏的人类细胞模型,无法在胚胎发育过程中进一步删除Zeb2依赖性调节网络,并且可以取消抗癌药物的发展。为此,我们使用CRISPR/CAS9介导的编辑系统生成了人类IPSC线,耗尽了Zeb2蛋白(表1)。我们分别应用了两个靶向Zeb2外显子5和外显子6的GRNA(图1 a),在父母IPSC线上Kicri002a(表1;(Uhlin等,2017)。通过LiPofection将包含两个GRNA的构建体引入IPSC系,并通过荧光激活的细胞分选(FACS)选择转染的细胞以表达绿色荧光蛋白。单细胞克隆在LN521上扩展,并通过基因组DNA上的Sanger测序分析基因编辑。分析显示了具有纯合790 bp缺失的克隆线kicri002a-4,跨越了内含子5和外显子5和6的一部分(chr2:g.144,404,077 - 144,404,404,404,867del;1 a;补充。图1 A-B)。 外显子5和外显子6的其余部分被融合,预测氨基酸194上的截短的Zeb2 mRNA,其截短的Zeb2 mRNA(PTC)(P.THR188888888888888888888888;图1 A-B)。外显子5和外显子6的其余部分被融合,预测氨基酸194上的截短的Zeb2 mRNA,其截短的Zeb2 mRNA(PTC)(P.THR188888888888888888888888;图1 a)。与136PTC位于编码N末端锌指(NZF)域的区域以及更C末端的R-SMAD结合域(SBD),CTBP相互作用结构域(CID)(CID)和C-末端的c-terminal Zinc Zinc Finger(CZF(CZF)和Homeododomain(例如Domains)(epifa)(epifa)。

Kalman滤波和期望最大化多阶段光谱unmixing
摘要 - 高光谱成像技术的最新演变和新的新兴应用程序的扩散按下了多个时间高光谱图像的处理。在这项工作中,我们提出了一种新型的频谱拆解(SU)策略,使用出色动机的参数末端记录来说明时间频谱变异性。通过使用状态空间公式来表示多个时空混合过程,我们能够利用贝叶斯过滤机制来估计末端的变异性系数。假设丰度的时间变化在短时间间隔很小,则采用了预期最大化(EM)算法的有效实施来估计丰度和其他模型pa-Rameters。仿真结果表明,所提出的策略优于最先进的多阶段算法。
标准PowerPoint P/Dis>
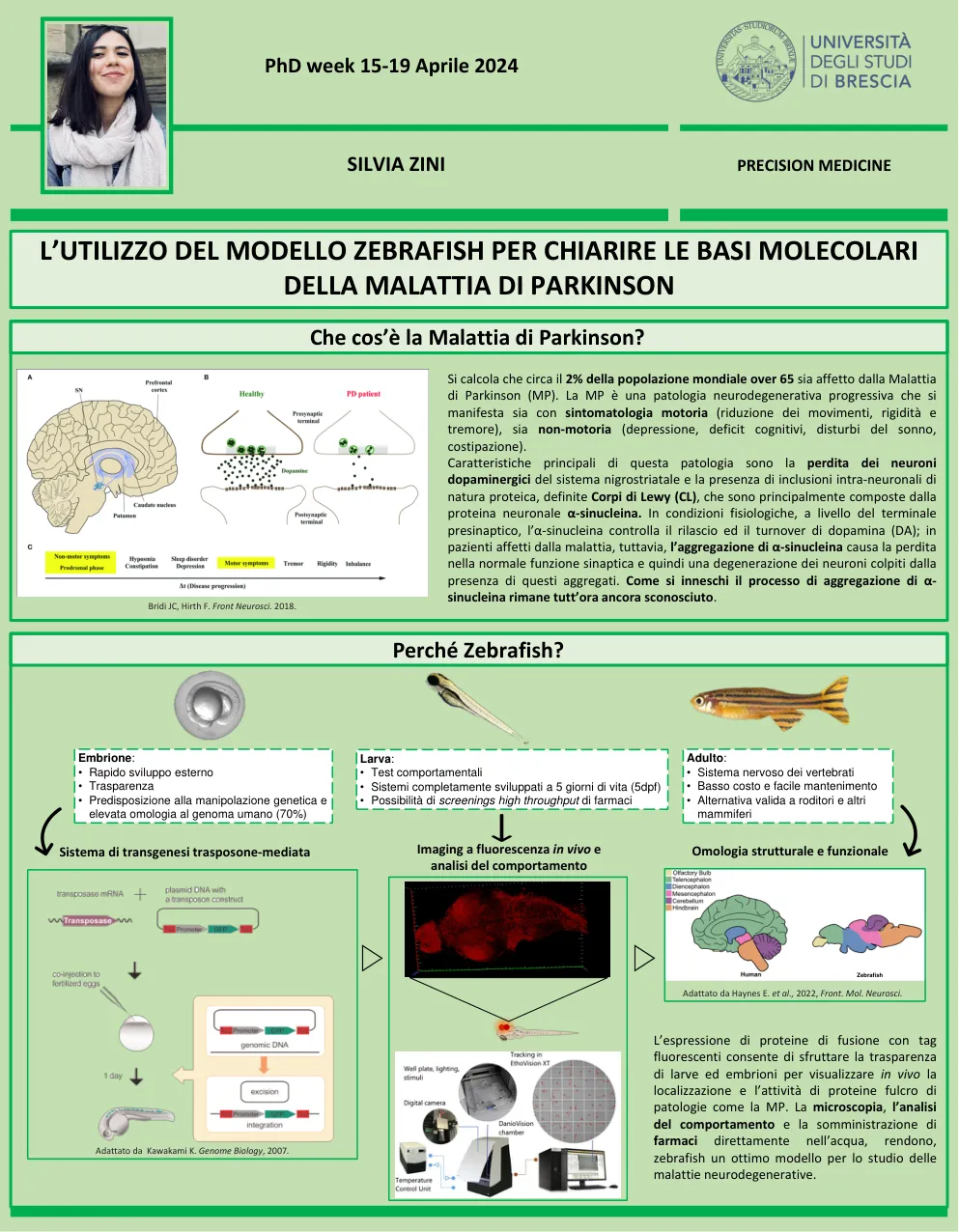
据估计,大约有65岁以上的世界人口中有2%患有帕金森氏病(MP)。MP是一种进行性神经退行性病理学,它既表现出运动症状(减少运动,刚性和震颤)和非运动(抑郁,认知缺陷,睡眠障碍,便秘)。这种病理学的主要特征是Nigrostria系统多巴胺能神经元的丧失以及蛋白质性质内神经内包含的存在,该蛋白质特征定义为Lewy(CL)体,主要由α-苏蛋白神经素神经元蛋白组成。在生理条件下,在pres虫末端的水平上,α-苏核素控制多巴胺的释放和周转(从);然而,在疾病患者中,α-苏核素的聚集会导致正常突触功能的丧失,因此会导致受这些聚集体影响的神经元的变性。如何触发α-苏核素的聚集过程仍然未知。
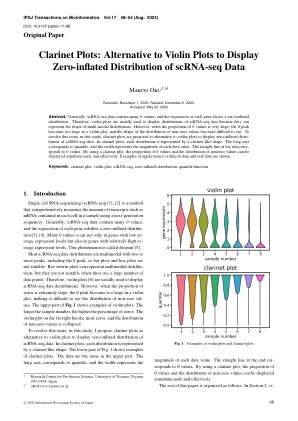
替代小提琴图以显示零充气...
摘要:通常,SCRNA-SEQ数据包含许多0值,每个基因的表达显示为零插入分布。因此,小提琴图通常用于显示SCRNA-Seq数据的分布,因为它们可以表示多模式分布的形状。但是,当0个值的比例非常大时,在小提琴图中,0峰变得太大,而非零值的分布形状很难看见。为了解决此问题,在这项研究中,提议了单簧管图作为小提琴图的替代方案,以显示SCRNA-SEQ数据的零空调。在单簧管图中,每个分布都以单簧管状的形状表示。长轴对应于分位数,宽度表示每个数据值的大小。末端的直线至0值。通过使用单簧管图,可以同时且有效地显示0个值的比例和非零值的分布。显示了用于人工数据的示例和实际数据。

人类染色体的起源2:祖先端粒 -
摘要我们已经从人类2,C8.1和C29B的两个等位基因组宇宙中鉴定出了两个等位基因组宇宙,每个粘液均包含两个脊椎动物端粒重复的倒置阵列,并在头对头排列,5'(ttaggg), - (ccctaa), - (ccctaa),3'。序列fln g这个端粒重复是当今人类序列的特征。BAL-31核酸酶实验人造人造染色体的克隆和荧光原位杂交的荧光表明,这些倒置重复的序列均与2 Q13和不同但重叠的人类染色体末端的子集杂交。我们得出的结论是,克隆在宇宙中C8.1和C29B中的基因座是古老的端粒融合的遗物,标志着两个祖先猿染色体融合产生人类染色体的点。