XiaoMi-AI文件搜索系统
World File Search System核质

kilodalton核帽结合蛋白
已经表明,单甲基化的帽结构在核事件中起着重要作用。盖结构与增强前mRNA剪接有关。最近,还建议这种结构促进RNA从细胞核到细胞质的转运。我们先前已经从HELA细胞核提取物中鉴定出并纯化了8OKD核盖结合蛋白(NCBP),这可能会介导这些核活性。在本报告中,我们描述了编码NCBP的互补DNA(cDNA)的克隆。确定了NCBP的部分蛋白质序列,并从HELA cDNA文库中分离出NCBP的全长cDNA。该cDNA编码了790个氨基酸的开放阅读框,其计算的分子质量为91,734 daltons,其中包含大多数确定的蛋白质序列。但是,蛋白质序列与任何已知蛋白质都没有显着同源性。转染实验表明,在HELA细胞中瞬时表达的表位标记的NCBP仅在核质中定位。使用截短的NCBP cDNA进行的类似实验表明,这种核定位活性由N末端70氨基酸区域赋予。

石墙后面:核脂质的热烈活动
生物分子的四种主要类型是核酸,蛋白质,碳水化合物和脂质。对他们各自互动的知识与对每个人的个人理解一样重要。然而,例如,对蛋白质与其他三组的相互作用进行了广泛的研究,但相比之下,核酸和脂质的相互作用探索了非常差。DNA和脂质之间的物理(且可能功能性)接近的标志性范式是真核生物中基因组DNA的情况:两个同心脂质双层构成核内的基因组DNA,这种相互作用的含义,这种相互作用的丰富,例如这种相互作用,例如,基因组稳定性,仍然是无关的。已经观察到了50年的核脂质相关表,但在大多数情况下,仅作为轶事描述。在这篇综述中,我们将汇总将脂质与核包膜和核质连接起来的证据,并将对这些描述进行批判性分析。我们的探索建立了一种场景,在这种情况下,脂质在核稳态中发挥了无可辩驳的作用。©2024作者。由Elsevier B.V.这是CC BY-NC-ND许可证(http://creativecommons.org/licenses/by-nc-nd/4.0/)下的开放访问文章。

氟对人类大脑发育的影响
氟化物会对正在发育的人脑产生有害的生化和功能变化。氟化物可能从母体血液中的氟通过胎盘传给胎儿开始。1-3 氟化物能穿过血脑屏障,在脑组织中蓄积的氟可能干扰脑磷脂的代谢,而这与神经元的退化有关。脑磷脂代谢的变化可能与慢性氟中毒的发病机制有关。我们对胎儿大脑的体视学研究显示,大脑皮层、海马锥体、浦肯野细胞和未分化神经母细胞的数值密度和核质比较高。但与非流行区相比,线粒体神经元的平均体积、数值密度和表面密度较低。根据 Rabinowich 的观点 5 ,神经元体积的数值密度增加和未分化神经母细胞是神经组织细胞形态不良的征兆。此外,细胞核-细胞质比增加反映了细胞增殖和成熟,蛋白质合成受到不利影响。在氟中毒大鼠中,RNA 损失会降低 ATP 的产生,从而导致代谢异常。6 综上所述,过量氟化物的这些影响反过来可能会促进血脑屏障的渗透,干扰 RNA 合成和酶促蛋白质代谢,并导致分化缓慢。

生物学二年级高中EJA细胞学核...
核是动物细胞的最大结构,并容纳染色体。每个染色体都包含几个基因,即指挥细胞活性的遗传物质。因此,我们说核心是遗传因素(从父母传播到子女)和细胞代谢活动的调节者。它是细胞的“重要中心”。核包络(腔室) - 围绕核内容物的膜,它具有许多孔,可以在核心和细胞质之间交换物质。通常,细胞活性越强烈,核中孔的数量越大。该膜存在于真核细胞中,但在原核生物中不存在 - 在这些细胞中,遗传材料直接浸入细胞质液中。核质(Cariolinfa) - 是填充细胞核内部空间的凝胶状材料。核仁 - 浸入cariolinfa中的圆形和非成膜小体。每个细丝都包含许多基因。在分裂细胞中,长而薄的染色质丝变短,更厚:然后被称为染色体。染色体负责遗传特征的传播,基本上由两种类型的化学物质形成:蛋白质和核酸。在染色体中发现的核酸是脱氧核糖核酸-DNA。

组合分子筛选确定了...
肥胖治疗对急性前临床白血病患者有效,体现了针对多种致癌调节成分的治疗的重要性。然而,最近的研究表明,急性髓样白血病(AML)的突变复杂性排除了分子靶向转化为临床成功的转化。在这里,作为基因分析的组合,我们使用了公正的,联合性的体外药物筛查来识别驱动AML并开发出良性的组合治疗的途径。首先,我们在原代AML细胞上筛选了513个自然量,并确定了一种新型的二萜(H4),该二萜(H4)优先诱导FLT3野生型AML的分化,而FLT3-ITD/突变赋予了抗性。对H4响应的样品显示出髓样标记的表达增加,核质质比的明显降低以及单核细胞转录程序重新激活的潜力减少了体内白血病的传播。通过使用H4和具有定义靶标的分子组合筛选,我们证明H4通过激活蛋白激酶C(PKC)信号通路的激活诱导分化,并激活PKC磷酸化和PKC到细胞膜的PKC磷酸化和易位。此外,组合筛选确定了溴和末端结构域(BET)抑制剂,该抑制剂可以进一步改善H4依赖性的白细胞分化在FLT3野生型单核细胞AML中。这些发现说明了用于开发AML组合治疗方法的公正,多重筛选平台的价值。

核进口缺陷驱动神经退行性中的细胞周期失调
神经退行性疾病(NDDS)和其他与年龄有关的疾病已通过一组关键的病理标志在经典上定义。这些标志中的两个,细胞周期失调(CCD)和核质转运(NCT)缺陷,长期以来一直在争论为因果关系,在加速衰老的病理学中是因果关系。具体而言,已证明有丝分裂后神经元中异常细胞周期活化会触发神经元细胞死亡途径和细胞衰老。此外,已经观察到NCT在衰老和神经变性过程中逐渐失调,其中增加了核蛋白的亚细胞再分配(例如TAR DNA-结合蛋白43(TDP43))对细胞质的主要驱动力是许多NDDS的主要驱动力。然而,NCT缺陷的功能意义是作为病理学的主要驱动因素或后果,以及细胞周期机械的重新分布如何促进神经变性,尚不清楚。在这里,我们描述了对进口素β进口的药理抑制能够在丝分裂神经元细胞系和有丝分裂后原发性神经元体外扰动细胞周期机制。以核进口缺陷为特征的运动神经元疾病的NEMF R86S小鼠模型,进一步概括了有丝分裂细胞系中CCD的标志,在体外和有丝分裂后的原发性神经元中以及体内脊柱运动神经元中。观察到的CCD与NDDS中神经元细胞死亡和细胞衰老中观察到的转录和表型失调一致。在一起,这些证据表明,导致CCD的核进口途径受损可能是神经变性中病理学的常见驱动力。

DNA 拓扑异构酶 IIα 剪接变体对获得性的影响...
摘要 DNA 拓扑异构酶 II α (170 kDa, TOP2 α /170) 诱导增殖细胞中瞬时 DNA 双链断裂,以解决染色体凝聚、复制和分离过程中的 DNA 拓扑纠缠。因此,TOP2 α /170 是抗癌药物的主要靶点,其临床疗效常常因化学耐药性而受到影响。尽管已经确定了许多耐药机制,但人类癌细胞系对 TOP2 α 界面抑制剂/毒药的获得性耐药通常与 Top2 α /170 表达水平的降低有关。我们实验室最近的研究,结合其他研究人员的早期发现,支持以下假设:对 TOP2 α 靶向药物的获得性耐药的主要机制是由于替代的 RNA 加工/剪接。具体而言,已报道了几种 TOP2 α mRNA 剪接变体,它们保留了内含子,并被翻译成缺乏核定位序列的截短 TOP2 α 异构体,随后导致核质分布失调。此外,内含子保留可能导致截短异构体缺乏核定位序列和活性位点酪氨酸 (Tyr805),而活性位点酪氨酸是形成酶-DNA 共价复合物所必需的,并且在存在 TOP2 α 靶向药物的情况下诱导 DNA 损伤。最终,这些截短的 TOP2 α 异构体导致药物对细胞核中的 TOP2 α 的活性降低并表现出耐药性。因此,对调节 TOP2 α 前 mRNA 的替代 RNA 加工的机制的完整表征可能会产生新的策略来规避获得性耐药性。此外,新型 TOP2 α 剪接变体和截短的 TOP2 α 同工型可用作药物耐药性、预后和/或直接未来 TOP2 α 靶向治疗的生物标志物。

工具用于定量分馏哺乳动物细胞的一般方法
引言真核核被核包膜(NE)包围,并以染色体形式包含大部分细胞的遗传物质(Clark等,2019; Webster等,2009)。它代表了几个膜 - 分隔的细胞器中最突出的,每个细胞器都有其自身和动态的组成(Cohen等,2018; Cole,2016; Schrader等,2015)。大分子的核质运输是在cy- toplasm和核之间发生的连续高度调节过程(Alberts等,2002; Christie等,2016; MacAra,2001; MacAra,2001; Silver,1991; Silver,1991; Wente and watee and Rout,2010; Yoneda; Yoneda》,1997年)。每种大分子的正确核细胞质定位是维持细胞稳态的关键(Bauer等,2015; Park等,2011)。分子进出核的转运是由嵌入在NE中的核孔复合物(NPC)所介绍的。单个NPC由约30种不同蛋白质的多个拷贝组成,称为核苷(Nups; Dultz等,2022; Stewart,2022; Tingey et al。,2022; Wing et al。,2022),并且不仅对核细胞的运输量不仅对核细胞的依恋; Al。,2022)。蛋白质的异常核细胞质定位已与许多人类疾病的发病机理有关,例如癌症,代谢,心血管和神经模型 - 生成性疾病(Chung等,2018; Holmes et al。有几种机制可能导致蛋白质错误定位,例如贩运机制的改变,蛋白质靶向信号的改变以及蛋白质的变化更具体地说,癌蛋白,抑制剂和其他与癌症相关的蛋白质的错误定位会干扰正常的细胞稳态,并导致肿瘤发育和转移(Wang and Li,2014)。

NSUN2 介导的 m5C 修改驱动替代......
原理:间变性甲状腺癌 (ATC) 是一种极具侵袭性的甲状腺癌,在初次诊断时经常表现为局部晚期浸润或远处转移,因此错过了手术干预的最佳窗口。因此,全身化疗和靶向治疗对于改善 ATC 的预后至关重要。然而,ATC 对常规治疗表现出显著的耐药性,这凸显了阐明这种耐药性背后的生物学机制并确定新的治疗靶点以克服它的必要性。方法:我们对来自 ATC 样本的大量和单细胞 RNA 测序 (scRNA-seq) 数据进行了全面分析,以筛选与多药耐药 (MDR) 相关的 m 5 C 修饰相关基因。然后,我们进行了 IC 50 测定、流式细胞术,并使用了 Nsun2 敲除的自发致瘤 ATC 小鼠模型来证明 NSUN2 促进了 ATC 中的 MDR。为了研究 NSUN2 介导的耐药机制,我们生成了 NSUN2 敲除的 ATC 细胞系并进行了转录组学、蛋白质组学和 MeRIP-seq 分析。此外,还进行了 RNA 测序和可变剪接分析以确定 NSUN2 敲除后的整体变化。我们通过糖蛋白染色、变性 IP 泛素化、核质分馏和 PCR 进一步探索了 NSUN2/SRSF6/UAP1 轴的潜在机制。最后,我们在体外和体内评估了小分子 NSUN2 抑制剂与抗癌药物的协同作用。结果:我们的研究结果表明,NSUN2 表达与 ATC 中的 MDR 显着相关。 NSUN2 充当 SRSF6 mRNA 上的 m 5 C 的“写入器”,ALYREF 充当 m 5 C 的“读取器”,从而诱导选择性剪接重编程并将 UAP1 基因的剪接形式从 AGX1 重定向到 AGX2。因此,AGX2 增强了 ABC 转运蛋白的 N 连接糖基化,通过防止泛素化介导的降解来稳定它们。此外,NSUN2 抑制剂可降低 NSUN2 酶活性并减少下游靶标表达,从而为克服 ATC 中的 MDR 提供了一种新颖且有希望的治疗方法。结论:这些发现表明 NSUN2/SRSF6/UAP1 信号轴在 ATC 的 MDR 中起着至关重要的作用,并将 NSUN2 确定为 ATC 化疗和靶向治疗的协同靶点。
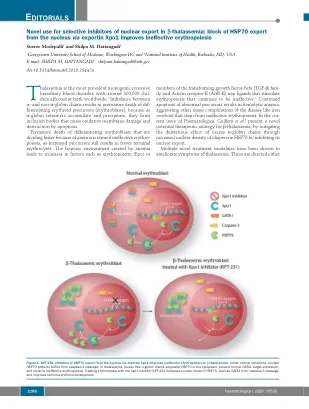
社论
改善未结合的A -Globin和非A -Globin链之间的平衡或纠正无效的红细胞。修饰的TFG-β家族受体拮抗剂,如Sotatercemp(ACE-011)和Luspatercept(ACE-536)阻止配体与ACTR-II受体结合,并随后激活SMAD4信号通路,4改善Ery-Throid Throid Cell和红细胞的产生。通过CRISPR Therapeatics成功的基因疗法实现了未结合A -Globin与非A -Globin链的异常比例,并得到了波士顿顶点药物的支持。称为CTX001的体细胞疗法使用了编辑的患者自己的造血干细胞(HSC)来刺激胎儿血红蛋白的产生。5通过XPO1抑制HSP70的细胞内局部局部局部局部可能会合并这两个治疗目标。几条证据表明,红细胞使用分子伴侣在红细胞发育过程中对不稳定的过量A -Glo- bin链分割,6-8,因此,靶向这种伴侣的靶向时,当过量的globobin tetramer会累积时,靶向这种伴侣可能在β -tha -thaplamasemia中有用。许多组指出,分子伴侣HSP70在红细胞9-11中积聚至高水平,对于简化胚芽成熟很重要。11正常的人红细胞成熟需要在成熟后期的caspase-3瞬时激活,以防止过度的红细胞生产。激活的胱天蛋白酶可以切割GATA-1,从而导致成熟停滞和/或凋亡。12 Ribeil等。 14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。12 Ribeil等。14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。表明EPO会导致HSP70转移到核中,结合GATA-1并保护其免受caspase-3裂解。相反,在EPO剥夺期间,HSP70被排除在细胞核中,而GATA-1被cas-pase-3裂解,导致凋亡死亡。13因此,HSP70的细胞内位置的改变似乎在红细胞生存力中起关键作用(图1)。在β-丘脑贫血中观察到的无效性红细胞生成的特征是在多染色体阶段加速了红细胞分化,成熟停滞和凋亡。在人β-thal虫蛋白粒细胞的成熟过程中,HSP70直接通过过量的游离A-格珠蛋白链直接在细胞质中螯合(图1)。核定靶向的HSP70突变体或caspase-3-无分解的GATA-1突变体恢复了β-thal核阿无血成红细胞的终末成熟。14在Haematologica,Guillem等。3跟进这种机制,以表明导出蛋白1(XPO1)调节在正常条件下HSP70在红细胞中的HSP70的核质质位置。Guillem等。证实,用XPO1抑制剂KPT-251治疗红细胞增加了HSP70的核水平,从caspase-3裂解中救出了GATA1,并改善了末端红细胞原理(图1)。尽管使用核出口的选择性抑制剂(SINE)用于治疗淋巴瘤和多骨髓瘤