XiaoMi-AI文件搜索系统
World File Search System核酶

新西兰泥浆蜗牛的基因组进化和渗入potamopyrgus estuarinus和potamopyrgus kaitunuparaoa
我们已经对Potamopyrgus estuarinus和Potamopyrgus kaitunuparaoa进行了测序,组装和分析的核和线粒体基因组和转录组,这是新西兰人的两个Prosobranch Snail物种,它们跨越了从河口到新淡水。这两个物种是淡水物种的最接近的亲属,potamopyrgus antipodarum是研究性别,宿主 - 寄生虫协同进化和生物侵入性的模型,因此为理解其异常生物学提供了关键的进化环境。P. esuarinus和P. kaitunuparaoa基因组的大小和整体基因含量非常相似。对基因组含量的比较分析,认为这两个物种具有涉及减数分裂和精子功能的几乎相同的基因,包括七个具有减数分裂特异性功能的基因。这些结果与这两个物种的强制性再生产是一致的,并为对抗杀虫假单胞菌的未来分析提供了一个框架,该物种既包含义务性的性和无性无性谱系,每个物种分别源自性祖先。全基因组多基因的系统发育分析表明,Kaitunuparaoa可能是最接近抗植物的。尽管如此,我们表明,埃斯图拉林和P. kaitunuparaoa之间存在相当大的渗透。该渗入不会扩展到线粒体基因组,该基因组似乎是雌雄同体和kaitunuparaoa P. estuarinus和P. kaitunuparaoa之间杂交的障碍。核编码基因,其产物在关节线粒体核酶复合物中的作用表现出相似的非渗透模式,这表明线粒体和核基因组之间的不兼容性可能阻止了这两种物种之间更广泛的基因流动。

一种用于裂殖酵母的快速有效的 CRISPR/Cas9 方法[...
摘要 CRISPR/Cas9 系统可实现无疤痕、无标记的基因组编辑。目前,用于裂殖酵母 Schizosaccharomyces pombe 的 CRISPR/Cas9 系统依赖于繁琐且耗时的克隆程序,将特定的 sgRNA 靶序列引入 Cas9 表达质粒中。此外,据报道,当从强 adh1 启动子持续过表达 Cas9 核酸内切酶时,它会对裂殖酵母产生毒性。为了克服这些问题,我们开发了一种改进的系统 SpEDIT,它使用从中等强度 adh15 启动子表达的针对 S. pombe 进行密码子优化的合成 Cas9 序列。SpEDIT 系统表现出灵活的模块化设计,其中 sgRNA 与自切割丁型肝炎病毒 (HDV) 核酶的 3' 端融合,从而允许 tRNA 基因序列中的 RNA 聚合酶 III 驱动 sgRNA 盒的表达。最后,在 GFP 占位符两侧加入 Bsa I 型 IIS 限制酶位点,可实现 Golden Gate 介导的一步式 GFP 替换和合成的 sgRNA 表达。SpEDIT 系统通过转化异步培养细胞,在 ade6 + 或 ura4 + 基因中生成靶点突变体,可实现 100% 的诱变效率。SpEDIT 还允许以最小的努力获得插入、标记和删除事件。还可以轻松实现两个独立非同源基因位点的同时编辑。重要的是,与目前可用的 S. pombe 编辑系统相比,SpEDIT 系统显示出更低的毒性。因此,SpEDIT 提供了一种有效且用户友好的 CRISPR/Cas9 程序,可显著改善裂殖酵母的基因组编辑工具箱。

基因组/基因编辑工具的观点
摘要:从理论上讲,可以区分等于或超过16 bp的DNA序列的DNA序列特异性识别蛋白可能是哺乳动物基因组独有的。长期序列的核酸酶,例如天然存在的归巢核酸酶和人工设计的ZFN,TALEN和CAS9-SGRNA。与其他对应物(通过蛋白质部分识别DNA靶位点的其他对应物相比,CAS9使用单个指南RNA(SGRNA)作为DNA靶标识别的模板。由于设计和合成目标位点的SGRNA的简单性,CAS9-SGRNA已成为基因组编辑的最新工具。此外,Cas9-SgrNA的RNA引导的DNA识别活性与HNH结构域和RUVC结构域的非平均链中的核酸酶活性无关,而HNH核酸酶无核酶无效无效无效活性无效(H 840 A)和RUVC核酸酶核酸酶活性无效null null突变(识别10 A)。与SGRNA,CAS9,Cas9(D 10 A),Cas9(H 840 A)和Cas9(D 10 A,H 840 A)一起用于实现双重链断裂,互补的链断管破裂,非满足链破裂,并且分别在TARPEC上进行破裂。基于这种独特的特征,可以在靶位点内或周围引入许多工程酶活性,例如DNA甲基化,组蛋白甲基化,组蛋白乙酰化,胞苷脱氨酸,腺嘌呤脱氨基和启动引导突变。为了防止Cas9衍生物的持久表达靶向,开发了许多瞬态表达方法,包括直接递送Cas9-SgrNA核糖蛋白。生物安全问题在体内应用中是必不可少的;已经设计了包装到病毒样颗粒或细胞外囊泡中的CAS9-SGRNA,已经报道了一些体内治疗试验。

有丝分裂原和应激激活蛋白激酶1负调节海马神经发生
摘要 - 成人海马的亚晶体区(SGZ)中的神经发生,可以通过多种手段来刺激,包括通过将实验动物暴露于丰富的环境中,从而提供额外的鼻子,社交和运动刺激。在丰富的动物中产生的有形健康和认知益处,包括改善对精神病,神经学和神经退行性疾病的建模,这可能会影响人类,这可能部分是由于神经元的产生增强所致。神经元反应富集的关键因素是释放脑衍生的神经营养因子(BDNF)和有丝分裂原活化蛋白激酶(MAPK)级联反应的激活,这可能导致刺激Neuroogenese或Neuroogenese的刺激。有丝分裂原和应激激活的蛋白激酶1(MSK1)是BDNF和MAPK下游的一种核酶,可调节转录。MSK1先前已经与缺乏MSK1蛋白的小鼠的研究有关基础和刺激的神经发生。在本研究中,使用仅缺乏MSK1激酶活性的小鼠,我们表明SGZ(KI-67染色)的细胞增殖速率没有由MSK1激酶DEAD(KD)突变造成的,并且与控制后水平的水平没有分歧。然而,与野生型小鼠相比,在标准housed和富集的MSK1 KD小鼠中,双铁蛋白(DCX)阳性细胞的数量都更大。2020年作者。由Elsevier Ltd代表IBRO出版。这是CC BY-NC-ND许可证(http://crea-tivecommons.org/licenses/by-nc-nd/4.0/)下的开放访问文章。这些观察结果表明,尽管MSK1不影响神经元前体的增殖基础速率,但MSK1负责调节注定成为神经元的细胞数量,可能是对新神经元数量的稳态控制,而新神经元的数量则是整合到齿状gyrus中的新神经元的数量。

速度编辑
摘要 CRISPR/Cas9 系统可实现无疤痕、无标记的基因组编辑。目前,用于裂殖酵母 Schizosaccharomyces pombe 的 CRISPR/Cas9 系统依赖于繁琐且耗时的克隆程序,将特定的 sgRNA 靶序列引入 Cas9 表达质粒中。此外,据报道,当从强 adh1 启动子持续过表达 Cas9 核酸内切酶时,它会对裂殖酵母产生毒性。为了克服这些问题,我们开发了一种改进的系统 SpEDIT,它使用从中等强度 adh15 启动子表达的针对 S. pombe 进行密码子优化的合成 Cas9 序列。SpEDIT 系统表现出灵活的模块化设计,其中 sgRNA 与自切割丁型肝炎病毒 (HDV) 核酶的 3' 端融合,从而允许 tRNA 基因序列中的 RNA 聚合酶 III 驱动 sgRNA 盒的表达。最后,在 GFP 占位符两侧加入 Bsa I 型 IIS 限制酶位点,可实现 Golden Gate 介导的一步式 GFP 替换和合成的 sgRNA 表达。SpEDIT 系统通过转化异步培养细胞,在 ade6 + 或 ura4 + 基因中生成靶点突变体,可实现 100% 的诱变效率。SpEDIT 还允许以最小的努力获得插入、标记和删除事件。还可以轻松实现两个独立非同源基因位点的同时编辑。重要的是,与目前可用的 S. pombe 编辑系统相比,SpEDIT 系统显示出更低的毒性。因此,SpEDIT 提供了一种有效且用户友好的 CRISPR/Cas9 程序,可显著改善裂殖酵母的基因组编辑工具箱。

长期暴露于多环芳烃污染物时,人皮肤菌群的变化
越来越多的证据表明,构成微生物组的人类肠道细菌与几种神经退行性疾病有关。在几项研究中发现了帕金森氏病(PD)和阿尔茨海默氏病(AD)患者的细菌种群的失衡。这种营养不良很可能会降低或增加分别具有保护性或有害人体的微生物组衍生的分子,并通过所谓的“肠脑轴”传达给大脑的这些变化。微生物组衍生的分子Queuine是一种富含大脑中的核酶,仅由细菌产生,并由人类通过其肠道上的表现来挽救。Queuine用枪支抗密码子在TRNA的Wobble位置(位置34)取代鸟嘌呤,并促进有效的细胞质和线粒体mRNA翻译。Queuine耗竭会导致蛋白质的折叠和激活,并激活小鼠和人类细胞中内质网应激和展开的蛋白质反应途径。蛋白质聚集和线粒体障碍通常与神经功能障碍和神经变性有关。为了阐明女王是否可以促进蛋白质折叠,并防止导致蛋白质病的聚集和线粒体缺陷,我们在几种化学合成的Queuine STL-101中测试了几种化学合成的女性STL-101的作用。用STL-101预处理神经元后,我们观察到高磷酸化的α-突触核蛋白的降低显着降低,α-突触核蛋白的标记是灰核核疗法的PD模型中α-突出蛋白聚集的标志物,并且在Accute and Actau consation and actau pyphosphoration中降低了Actuce and Actau phossephose contau pysease contau pysepy pd。此外,在AD模型以及PD的神经毒性模型中,在用STL-101预处理的细胞中发现了神经元存活的相关增加。测量180个神经健康个体血浆中的queuine表明健康的人类维持皇后区的保护水平。我们的工作已经确定了女性在神经保护中的新作用,从而发现了神经系统疾病中STL-101的治疗潜力。

可重新配置的二维DNA晶格
2纽约大学化学系,纽约,纽约10003,美国 *通讯作者。电子邮件:bw@tsinghua.edu.cn(B.W.); ned.seeman@nyu.edu(n.c.s.)。抽象的分支DNA基序是所有合成DNA纳米结构的基本结构元素。但是,分支方向的精确控制仍然是进一步增强整体结构秩序的关键挑战。在这项研究中,我们使用两种策略来控制分支方向。第一个基于固定的霍利迪连接,该连接在分支点上采用特定的核苷酸序列,以决定其方向。第二个策略是使用角度构造支柱在分支点上使用柔性垫片固定分支方向。我们还证明,可以通过规范的Watson-Crick碱基配对或非典型的核酶相互作用(例如I-MoTIF和G-Quadruplex)动态地实现分支方向控制。具有从化学环境的精确角度控制和反馈,这些结果将使新型的DNA纳米力学传感设备和精确有序的三维体系结构。在过去的四十年中,随着DNA纳米技术的快速发展,多功能的DNA纳米结构具有越来越增强的复杂性[1] [1]。作为分支结构基序在DNA纳米结构中无处不在,对螺旋分支的精确角度控制是关键挑战之一。相比之下,几何控制在很大程度上避开了DNA网络设计。对这些方案的拓扑控制已在很大程度上通过序列设计,螺旋时期和连接连通性的处方[2]阐明。Angle and lattice morphology is generally observed to be an emergent property of topological self-assembly—indeed the tensegrity triangle, a hallmark three-dimensional (3D) DNA lattice [3] , has three attainable internal angles, 101 º, 111 º, and 117 º, which is an apparent result of lattice stress by changing the edge length in otherwise topologically-similar structures.考虑到这一点,在现场中,获得更高的结构顺序(包括拓扑和几何特性)仍然是一个关键的挑战,可以作为实现设计师纳米材料功能的更雄心勃勃的目标的基础(例如酶促活动,刚性晶体支架,固定的晶体支架,纳米粒子阵列等)。类似于减数分裂的移动霍利迪交界处的固定的四臂连接是DNA纳米技术中最早的结构图案[2A,4]。它不仅在由无脚手架的DNA“乐高”方法构建的纳米结构中广泛使用[5],而且还使用脚手架的DNA折纸方法在不同的结构中呈现[6]。已证明分支方向由分支点序列[7]和交叉类型[8]定义,这表明了精确几何控制的机会。这种合成性指出了具有精确和动态原子布置的高阶DNA纳米结构的可行性。

配方,开发,表征和体内抗...
摘要本研究的目的是开发hesperidin植物体的配方,表征和体内抗糖尿病评估。使用卵磷脂45毫克制备制剂,精确称重的胆固醇15 mg,将其溶解在10 mL氯仿中,在圆底烧瓶(RBF)中,并进行10分钟的浴室超声处理。使用旋转蒸发器将有机溶剂除去45-50摄氏度。完全去除溶剂后形成的磷脂混合物薄层。Hesperidine旋转蒸发器用于在37-40°C下进行一小时的水合。透射电子显微镜用于检查植物体的形态。被应用于400个网状碳涂层的铜网格后,使用1%W/V磷酸烟酸对植物体分散剂进行负染色。使用Malvern Mastersizer S Laser衍射尺寸分析仪(Malvern Instruments Ltd.,UK)检查植物体的尺寸分布。使用文献中先前描述的方法,评估了体内抗糖尿病活性。Wistar大鼠,并将其保存在动物屋设施中,并带有12小时的浅色和黑暗周期。使用自动异性腔中的诊断试剂盒(ERBA诊断曼海姆,德国)用于估计生化参数。选择F1和F2批次作为最佳配方,然后根据形态(数字照片和TEM),粒径和封装效率进行进一步评估。囊泡范围从100 nm到500 nm不等。F1和F2植物体的平均大小分别为109.71和133.24 nm。在某些地区,胰岛和腺泡细胞(外分泌组织)之间的外围扩大较小。现在,两个单元都彼此接近,表明恢复正常。总而言之,基于植物体的公式可能是提高治疗功效,较低剂量和增强剂量方案的有用策略。为了要求其抗糖尿病特性,必须确认更多涉及人类受试者的研究。关键字:配方,表征,体内,抗糖尿病评估,hesperidin,植物体如何引用本文:Borkar S,Swapnil Goyal。配方,发育,表征和体内抗糖尿病植物体的抗糖尿病评估。国际药物输送技术杂志。2024; 14(4):2244-48 doi:10.25258/ijddt.14.4.41支持来源:nil。利益冲突:无引入,而“有些”是指类似细胞的,“ phyto”是指植物。1植物体是囊泡药物输送系统,可改善低溶剂的药物吸收和生物可利用性。1,2植物提取物和磷脂酰胆碱(或任何亲水极性头组)对形成植物体反应,它们是磷脂的复合物,并且天然存在的活性植物化学物质结合在其结构中。3,4与常见制剂相比,这些配方显示出更好的药理和药代动力学特征。亲水性植物核酶 - 胆碱络合物完全被脂溶性磷脂酰基部分覆盖。),例如多酚。高药物封装,更好的稳定性(在两亲分子的植物构成和极性头部之间形成化学键,5和改善的生物利用度6只是植物体的令人印象深刻的优势。唯一可以掺入植物体结构的植物化学物质是包含活性氢原子(-COOH,-OH,-NH2,-NH等)的植物化学物体。两亲分子的亲水部分和草药衍生物可以建立与

Rudeus,一种用于研究DNA结合蛋白的机器学习分类系统
工程大肠杆菌菌株用于生产长的单链DNA Konlin Shen 1,Jake J.洪水2,Zhuizi Zhang 1,Alvin HA 4,5,6,Brian R. Shy 4,5,6,John E.美国加利福尼亚州伯克利的国家实验室4美国加利福尼亚大学旧金山分校,美国加利福尼亚州旧金山的实验室医学系。5 Gladstone-UCSF基因组免疫学研究所,美国加利福尼亚州旧金山。6加利福尼亚大学旧金山分校的医学系,美国加利福尼亚州旧金山。 对应证:shawn.douglas@ucsf.edu抽象的长单链DNA(SSDNA)是一种多功能分子试剂,其应用包括RNA引导的基因组工程和DNA纳米技术,但其生产通常是资源密集的。 我们采用了一种新的方法,利用工程化的大肠杆菌“助手”菌株和吞噬系统,将长ssDNA的产生简化为直接转化和纯化程序。 我们的方法通过将M13MP18基因直接整合到大肠杆菌染色体中,从而消除了对辅助质粒及其相关污染的需求。 ,我们实现了504至20,724个核苷酸的ssDNA长度,碱性赖氨酸溶液纯化后滴度最高为250 µg/l。 通过将其在原代T细胞基因组修饰和DNA折纸折叠中的应用中,我们的系统的功效得到了证实。6加利福尼亚大学旧金山分校的医学系,美国加利福尼亚州旧金山。对应证:shawn.douglas@ucsf.edu抽象的长单链DNA(SSDNA)是一种多功能分子试剂,其应用包括RNA引导的基因组工程和DNA纳米技术,但其生产通常是资源密集的。我们采用了一种新的方法,利用工程化的大肠杆菌“助手”菌株和吞噬系统,将长ssDNA的产生简化为直接转化和纯化程序。我们的方法通过将M13MP18基因直接整合到大肠杆菌染色体中,从而消除了对辅助质粒及其相关污染的需求。,我们实现了504至20,724个核苷酸的ssDNA长度,碱性赖氨酸溶液纯化后滴度最高为250 µg/l。通过将其在原代T细胞基因组修饰和DNA折纸折叠中的应用中,我们的系统的功效得到了证实。我们的方法的可靠性,可伸缩性和易度性有望解锁需要大量长ssDNA的新实验应用。引言单链DNA(ssDNA)在生物技术中起着至关重要的作用,尤其是在DNA纳米技术和基因编辑1,2中。长ssDNA的合成超过5000个核苷酸(NT)是具有挑战性的,并且明显的障碍可以阻止可扩展产生。通过磷酰胺化学的直接化学合成仅限于由于掺入误差和脱尿3的长度300-400 nt。为了获得更长的ssDNA链,电流实践采用双链DNA(dsDNA)作为模板。例如,不对称PCR可以在长度4中产生高达15,000 nt的ssDNA。其他方法包括使用差异修饰的引物进行PCR扩增:用于lambda外核酶消化5的磷酸化和未磷酸化,或生物素基化和非生物素化和非生物素化,用于链霉亲和素珠分离6-在孤立的Ssdna strands隔离时进行抗性。然而,这些技术通常每50-微晶(µL)反应产生小于1微克(µg)的ssDNA,从而使毫克的生产量成本昂贵,并且由于广泛的劳动力和高度试剂的消耗而效率低下,因此强调了更多可扩展和经济的SSDNA生产方法的必要性。
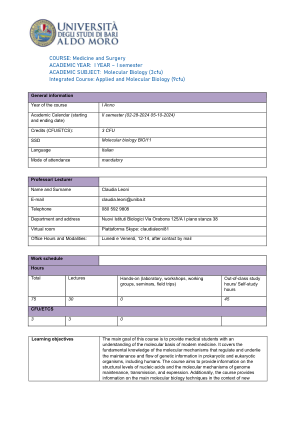
分子生物学(3cfu)综合课程
超螺旋和拓扑性质。拓扑异构酶。细菌类核。组蛋白和核小体的性质和组装。染色质的高级结构。组蛋白的翻译后修饰。溴多胺和染色质结构域。表观遗传学。原核生物和真核生物的基因组。复制模型。DNA合成。细菌DNA聚合酶。校对和缺口翻译。复制子模型。OriC和半甲基化。Ter/Tus。真核细胞核中的复制工厂。ARS结构和复制控制。酶学。前RC和前启动复合物。复制抑制剂,如化疗药物和抗病毒药物。端粒和端粒酶的结构、功能和意义。DNA损伤和修复。基因组作为动态实体。体细胞和种系突变。SNP。内在和外在损伤。化学和物理诱变剂。原核生物和真核生物中的去除、逆转和损伤避免系统。MUT 系统。BER 系统。糖基化酶的重要性。安全系统。NER 系统:UvrABCD 和 XP 蛋白。GG-NER 和 TC-NER。光解作用、MGMT、AlkBH。损伤耐受机制。TLS。细菌中的 SOS 反应。单丝和双丝断裂。HR 和 NHEJ。由于修复系统突变而导致的人类疾病。位点特异性重组。重组酶。Lambda 噬菌体。Cre-Lox 系统和 KO 小鼠。简单和复杂的转座子。SINE 和 LINE 元素、Alu 序列。原核生物和真核生物中的 RNA。结构、类型和特性。细菌 RNA 聚合酶和相关因子。转录单位。转录步骤。细菌启动子中的共识序列。终止机制。抑制剂。 Lac、ara 和 trp 操纵子。阳性和阴性对照。真核细胞中的 RNA 类别。RNA 聚合酶 (CTD) 的结构和功能。三种启动子的特征。基础转录机制。TFIIH。反式激活因子、辅激活因子。CpG 岛甲基化。组蛋白密码。长程调节剂。DNA 结合蛋白的功能域 (HTH、HD、HLH、ZF、LZ)。RNA 成熟、核运输和转录后控制。加帽类型。添加 polyA。CTD 的变化。外显子和内含子。外显子改组。四类内含子及其去除机制。剪接体和剪接位点。AT-AC 剪接。EJC 复合体。可变剪接。ESE 和 ESS 序列、SR 和 hnRNP 蛋白。SMN 基因。剪接和病理。rRNA 和 tRNA 加工反应。核糖体基因。 SnoRNA 和核仁功能。RNA 编辑。插入和转换编辑。人类 RNA 编辑的示例。细胞核和细胞质中的 RNA 周转。外泌体。无义介导的 mRNA 衰变 (NMD)。非编码 RNA。小 RNA 在细胞中的功能。RNA 干扰。siRNA。微小 RNA 的生物发生。miRNA、长链非编码 RNA、环状 RNA 的作用机制。逆转录病毒的一般信息。遗传密码和翻译。遗传密码的性质和特征。线粒体密码。ORF。tRNA 的特征。不常见碱基。aa-tRNA 合成酶的功能和类别。遗传密码的翻译重编码和扩展。SeCys。核糖体是一种核酶。原核生物和真核生物的翻译阶段。不同的启动机制。能量成本。NSMD。细菌中的 tmRNA。抑制剂。蛋白质的翻译后修饰、分选和降解。折叠和错误折叠。朊病毒。HSP60 和 HSP70。泛素和泛素化系统。SUMO 化糖基化。蛋白酶体。肽信号。蛋白质分选。线粒体输入。线粒体基因组细胞中的线粒体可塑性。人类线粒体基因组。遗传、结构、复制及其表达的原理。线粒体 DNA 中的改变。DNA 克隆的原理。修饰限制系统。克隆载体。cDNA 合成。基因组 DNA 和 cDNA 文库。TA 克隆。表达克隆。基因表达沉默。基因治疗。数据库。基因组编辑元件(Talen、Zn 指、CRISPR/Cas9 系统)。PCR 和 DNA 测序。PCR 的特性。PCR-RFLP。实时 PCR、DNA 测序。NGS。核酸杂交。杂交原理。熔点和严格性。探针制备:切口平移。Southern、Northern、杂交测定。蛋白质印迹。