XiaoMi-AI文件搜索系统
World File Search System氟乙酸
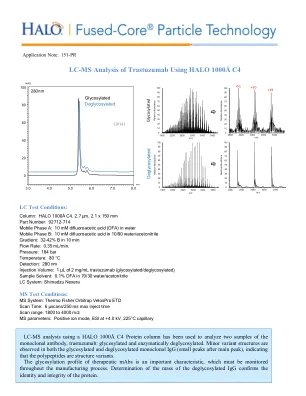
使用 HALO 1000Å C4 对曲妥珠单抗进行 LC-MS 分析
色谱柱:HALO 1000Å C4, 2.7 µm, 2.1 x 150 mm 部件号:92712-714 流动相 A:10 mM 二氟乙酸 (DFA) 水溶液 流动相 B:10/90 水/乙腈中的 10 mM 二氟乙酸 梯度:10 分钟内 B 从 32% 变为 42% 流速:0.35 mL/min。压力:184 bar 温度:80 °C 检测:280 nm 进样量:1 µL 2 mg/mL 曲妥珠单抗(糖基化/去糖基化) 样品溶剂:0.1% DFA 溶于 70/30 水/乙腈 LC 系统:Shimadzu Nexera MS 测试条件: MS 系统:Thermo Fisher Orbitrap VelosPro ETD 扫描时间:6 µscans/250 ms 最大进样时间 扫描范围:1800 至 4000 m/z MS 参数:正离子模式,ESI 在 +4.0 kV,225°C 毛细管

食品包装应用的透明,增塑的纤维素 - 甘油生物塑料
通过滴脂糖甘油混合物(高达50 wt%甘油)溶解在三氟乙酸和三氟乙酸酸酐(TFA:TFA:TFAA:TFAA,2:1,2:1,V:V:v)中,获得了自由膜。进行了膜的光学,结构,机械,热力学,屏障,迁移,防油性和生物降解特征的全面检查。所得的纤维素 - 甘油混合物分别表现出无定形分子结构和增强的H键网络,分别通过X射线衍射分析和红外光谱证明。包含甘油对膜的机械性能产生了塑性影响,同时保持其透明度。通过水吸收和水蒸气/氧气传输速率评估流体动力和屏障性能,并且获得的值与其他基于纤维素的材料的值一致。此外,总体迁移水平低于欧盟的调节限制,如使用Tenax®作为干粮模拟剂所述。此外,这些生物塑料表现出良好的防油性性能,尤其是在高甘油含量的情况下,以及作为烘焙产品包装材料的潜力。通过测量海水中的生物氧需求,观察到甘油诱导的高生物降解率,进行了生物降解性评估。


通过单个直接注入LC-MS/MS方法
每氯烷基酸(PFAAS),例如三氟乙酸(TFA),氟丙烷酸(PFPRA),丙烷磺酸(PFMS),丙酸(PFMS),丙烷基硫酸硫酸硫酸硫酸(PFROROUR)(PFROROUR)(PFROROUD), PFA的一个子集,其特征是每氟化碳(C F)的链长度为1-3。 1与它们的长链对应物相比,这些化学物质在历史上被忽略了,原因是它们的毒性较低和生物蓄积潜力。 然而,这些超短链PFAA的高极性,水溶性和持久性会导致在水生和植物环境中积累,从而增加水生生物和人类的暴露。 尤其是在全球范围内报道了TFA在水性,固体和生物矩阵中的报道,通常比长链PFAA的浓度高。 2除了直接来源(例如工业生产)外,TFA还据报道是流通制冷剂,农药和药物的降解产物。 3,4这些正在进行的排放,加上TFA的极端持久性和流动性,导致了迅速增加和潜在不可逆转的行星暴露。 2每氯烷基酸(PFAAS),例如三氟乙酸(TFA),氟丙烷酸(PFPRA),丙烷磺酸(PFMS),丙酸(PFMS),丙烷基硫酸硫酸硫酸硫酸(PFROROUR)(PFROROUR)(PFROROUD), PFA的一个子集,其特征是每氟化碳(C F)的链长度为1-3。1与它们的长链对应物相比,这些化学物质在历史上被忽略了,原因是它们的毒性较低和生物蓄积潜力。然而,这些超短链PFAA的高极性,水溶性和持久性会导致在水生和植物环境中积累,从而增加水生生物和人类的暴露。尤其是在全球范围内报道了TFA在水性,固体和生物矩阵中的报道,通常比长链PFAA的浓度高。2除了直接来源(例如工业生产)外,TFA还据报道是流通制冷剂,农药和药物的降解产物。3,4这些正在进行的排放,加上TFA的极端持久性和流动性,导致了迅速增加和潜在不可逆转的行星暴露。2

疏水素:多任务蛋白
疏水性是由纤维真菌产生的小两亲性细胞外蛋白。它们是表面活性蛋白,它们的功能主要与它们在疏水 - 亲水性接口处自我组装成两亲性单层的能力有关。取决于其水文模式和纯粹的要求,它们被分为I类和II类;两者都在整个序列中均表现出八个保守的半胱氨酸,形成了四个拆桥,它们产生了四个循环,可以使蛋白质以其单体和折叠形式稳定。I类杂菌环比II类杂菌环更扩展,从而导致不同表面的组装差异,并伴随着蛋白质结构的构象变化。 在单体杂素糖基化形式中,疏水素富含β-地表结构,同时在水中组装时 - 空气界面在其结构中增加了β-单表的含量,并且与水的界面和疏水固体在界面上,以及诸如TE的杂化固体,例如TE的形成也诱导了α-α-α-α-α-α-α-elix -Helix -Helix -Helix -Helix -Helix -Helix -Helix -a -Helix -a -Helix -α-固定结构。 由I类生成的单层是稳定的结构,称为纤维或rodlets,II类仅产生聚集体。 I类在其序列中呈现糖基化链。这会导致α-螺旋结构的形成,从而促进有序的组件,这需要它们的稳定性和高不溶性。 原纤维可以与三氟乙酸和甲酸分离,而三乙酸可以展开蛋白质,而60%乙醇和2%十二烷基硫酸钠溶液解散了II类聚集体。I类杂菌环比II类杂菌环更扩展,从而导致不同表面的组装差异,并伴随着蛋白质结构的构象变化。在单体杂素糖基化形式中,疏水素富含β-地表结构,同时在水中组装时 - 空气界面在其结构中增加了β-单表的含量,并且与水的界面和疏水固体在界面上,以及诸如TE的杂化固体,例如TE的形成也诱导了α-α-α-α-α-α-α-elix -Helix -Helix -Helix -Helix -Helix -Helix -Helix -a -Helix -a -Helix -α-固定结构。由I类生成的单层是稳定的结构,称为纤维或rodlets,II类仅产生聚集体。I类在其序列中呈现糖基化链。这会导致α-螺旋结构的形成,从而促进有序的组件,这需要它们的稳定性和高不溶性。 原纤维可以与三氟乙酸和甲酸分离,而三乙酸可以展开蛋白质,而60%乙醇和2%十二烷基硫酸钠溶液解散了II类聚集体。I类在其序列中呈现糖基化链。这会导致α-螺旋结构的形成,从而促进有序的组件,这需要它们的稳定性和高不溶性。原纤维可以与三氟乙酸和甲酸分离,而三乙酸可以展开蛋白质,而60%乙醇和2%十二烷基硫酸钠溶液解散了II类聚集体。

使用镍(II)Allyl系统对丁二烯的协调链转移聚合:直接径丁二烯的直接途径
摘要:镍烯丙基复合物是丁二烯(BD)1,4-会员聚合物的催化剂。协调链转移聚合(CCTP)尚未使用这些系统评估。我们在这项工作中报告了丁二烯在存在π-甲基镍(II)三氟乙酸(TFA)和MG N BUET或ALET 3作为链转移剂(CTA)案例研究的情况下的聚合。反应遵循一阶动力学与单体相比。在CTA存在的情况下证明了链的转移,并形成带有共轭二烯部分的多丁二烯。这允许通过重新插入链条一锅访问分支多丁二烯。多丁二烯氢化后,通过13 c NMR定量分析分支,并评估了其对氢化样品的热性能的影响,特别是对于无法定量确定的低度分支。暂时提供了催化循环的完整描述。如果在乙烯聚合过程中在文献中描述了类似的串联过程,据我们所知,这是迄今为止报道的唯一用于共轭二烯的系统,导致分支多丁二烯,从而扩展了CCTP过程的应用范围。■简介

n- ...
摘要:已被广泛接受的是,诸如HCl之类的酸性物种抑制了N-羧基酸酐(NCA)的聚合过程,必须将其去除以保证成功合成多肽。在这里,我们表明有机酸对NCA聚合的影响取决于其在二氯甲烷中的PKA值。虽然较强的酸(例如三氟乙酸)完全阻止了链的传播,但较弱的酸(例如乙酸)会加速聚合速率。酸的添加不仅质子化了传播的氨基群,还激活了NCA单体,其平衡确定了催化作用或抑制作用。此外,酸催化的聚合表现出与常规合作共价聚合物不同的一阶段动力学,即使加速速率也可以很好地控制分子量。PKA依赖性促使我们按需将抑制酸 - 将抑制作用转化为催化剂,从而促进了来自非纯化NCA单体的受控聚合。这项工作强调了通过改变反应条件来改变对催化剂/抑制剂的常规理解的可能性,这不仅阐明了新催化剂的设计,而且还提供了一种实用策略,以有效和控制的方式准备多肽材料。

设计纳米结构核仁素结合肽用于三阴性乳腺癌干细胞内的细胞内药物输送
101 肽合成。使用 2-氯三苯甲基氯树脂,按照 104 Fmoc/t Bu 合成策略,手工合成 103 hv6pep、pw14、sCH9 和 p17 肽配体的羧基荧光素衍生物。使用二异丙基碳二酰亚胺 (DIC) 和 HOBt · H 2 O 作为偶联剂 105 进行肽延伸,并通过用 107 哌啶/DMF(2:8,v/v)处理进行 Fmoc 消除。肽序列延伸 108 完成后,将每个肽基树脂分成两部分。一部分 109 用直接连接到肽配体序列 N- 110 末端的羧基荧光素 (CF) 衍生化。另一部分,将 Fmoc-6-氨基己酸间隔基引入肽序列的 N 端,随后在先前消除 Fmoc 保护基后用 CF 进行修饰。通过用三氟乙酸 (TFA/H 2 O/TIS,95:2.5:2.5) 进行酸解处理,将肽从树脂上切下。采用收敛策略,使用两个受保护的肽片段 (片段 F3A (1-15) 和片段 F3B (16-30)) 合成了羧基荧光素化的肽配体 F3。两个肽片段均在 Liberty Lite 微波炉上合成

封闭的芳香管——Capsularenes - UCL Discovery
摘要:在本研究中,我们描述了一种将芳烃掺入封闭管(我们将其命名为胶囊烯)的合成方法。首先,我们制备了花瓶状的分子篮 4 – 7 。这些分子篮由一个苯碱基和三个双环[2.2.1]庚烷环融合而成,这些环延伸到邻苯二甲酰亚胺 ( 4 )、萘二甲酰亚胺 ( 6 ) 和蒽二酰亚胺侧 ( 7 ),每个侧都带有一个二甲氧基乙烷缩醛基团。在催化三氟乙酸 (TFA) 的存在下,4、6 和 7 顶部的缩醛转变为脂肪族醛,随后在分子内环化为 1,3,5-三氧杂环己烷(1 H NMR 光谱)。这种环闭合几乎是一个定量过程,它提供了不同大小的胶囊烯 1 (0.7×0.9 纳米)、8 (0.7×1.1 纳米;) 和 9 (0.7×1.4 纳米;),这些胶囊烯的特征是 X 射线晶体学、微晶电子衍射、紫外/可见光、荧光、循环伏安法和热重法。胶囊烯具有出色的刚性、独特的拓扑结构、出色的热稳定性以及可能可调的光电特性,有望用于构建新型有机电子设备。

1 EnzyHTP 计算定向进化与...
摘要:定向进化通过迭代诱变促进酶工程。尽管高通量筛选应用广泛,但构建“智能库”以有效识别有益变体仍然是该社区面临的主要挑战。在这里,我们基于 EnzyHTP 开发了一种新的计算定向进化协议,EnzyHTP 是我们之前报道过的用于自动化酶建模的软件。为了提高吞吐效率,我们实施了一种自适应资源分配策略,该策略根据工作流中酶建模子任务的特定需求动态分配不同类型的计算资源(例如 GPU/CPU)。我们将该策略实现为 Python 库,并使用氟乙酸脱卤酶作为模型酶测试了该库。结果表明,与在整个工作流中 CPU 和 GPU 都随时可用的固定资源分配相比,应用自适应资源分配可以节省 87% 的 CPU 小时数和 14% 的 GPU 小时数。此外,我们在自适应资源分配框架下构建了一个计算定向进化协议。该工作流程在 Kemp 消除酶定向进化实验中针对两轮突变筛选进行了测试,总共有 184 个突变体。使用折叠稳定性和静电稳定能作为计算读数,我们重现了四个实验观察到的目标变体中的三个。借助该工作流程,整个计算任务(即 18.4 μs MD 和 18,400 QM 单点计算)在三天内使用约 30 个 GPU 和约 1000 个 CPU 完成。