XiaoMi-AI文件搜索系统
World File Search System氟勃罗病

克罗恩病靶向治疗续保
请将请求传真至 1-800-609-4884(免费)或将请求邮寄至:PharmaCare, Box 9652 Stn Prov Govt, Victoria, BC V8W 9P4 此传真件为医患专用,包含仅供 PharmaCare 使用的机密信息。严禁任何其他分发、复印或披露。如果 PharmaCare 批准此特别授权请求,则批准仅用于支付处方费用。PharmaCare 批准并不表明所请求的药物是否适合任何特定患者或病症。缺少信息的表格将被退回以供填写。如果没有提供处方者传真或邮寄地址,PharmaCare 将无法回复。

ZORYVE(罗氟司特)乳膏政策 I. 适应症...
1. 会员年满 12 岁,被诊断患有斑块状银屑病 2. 受影响的 BSA 介于 2-20% 之间。提供受影响的 BSA 的证明文件。 3. Zoryve 由皮肤病学或风湿病学专家开具处方或咨询。 4. 会员在至少连续 2-4 周的试验中治疗反应不足

小胶质细胞TREM2调控阿尔茨海默病发生的机制研究进展
Engl J Med,2013,368:107-16 [4] Jin SC,Benitez BA,Karch CM等。trem2中的编码变体增加了阿尔茨海默氏病的风险。Hum Mol Genet,2014,23:5838-46 [5] Schwabe T,Srinivasan K,Rhinn H.移动范式:小胶质细胞在阿尔茨海默氏病中的核心作用。Neurobiol Dis,2020,143:104962 [6] Zhang Y,Chen K,Sloan SA等。大脑皮层的神经胶质,神经元和血管细胞的RNA测序转录组和剪接数据库。J Neurosci,2014,34:11929-47 [7] Lloyd AF,Miron Ve。小胶质细胞在中枢神经系统中的促估计性特性。nat Rev Neurol,2019,15:447-58 [8] Butovsky O,Ziv Y,Schwartz A等。由IL-4或IFN-γ激活的小胶质细胞差异地诱导了成人茎/祖细胞的神经发生和寡构成。mol Cell Neurosci,2006,31:149-60 [9] Ulland TK,Song Wm,Huang SC等。TREM2在阿尔茨海默氏病中保持小胶质细胞代谢适应性。Cell,2017,170:649-63.E13 [10] Daws MR,Lanier LL,Seaman WE等。新型小鼠髓样DAP12-相关受体家族的克隆和表征。EUR J Immunol,2001,31:783-91 [11] Dean HB,Roberson ED,Song Y. Trem2中与神经退行性疾病相关的变体破坏了免疫球蛋白领域的顶端配体结合区域。前神经,2019,10:1252-67 [12] Sasaki A,Kakita A,Yoshida K等。小胶质细胞DAP12和TREM2基因在NASU-Hakola病中的可变表达。神经遗传学,2015,16:265-76 [13] Jay TR,Von Saucken VE,Landreth GE。trem2在神经退行性疾病中。mol Neurodegener,2017,12:56-89 [14] Forabosco P,Ramasamy A,Trabzuni D等。通过人脑基因表达数据网络分析对TREM2生物学的见解。Neurobiol老化,2013,34:2699-714 [15] Schlepckow K,Kleinberger G,Fukumori A等。与阿尔茨海默氏症相关的trem2变体发生在亚当裂解位点,并效果脱落和吞噬功能。embo mol Med,2017,9:1356-65 [16] Bouchon A,Dietrich J,ColonnaM。尖锐边缘:炎症反应可以由Trem-1触发,Trem-1是一种在中性粒细胞和单核细胞上表达的新型受体。J Immunol,2000,164:4991-5 [17] Del-Aguila JL,Benitez BA,Li Z等。 TREM2脑转录本特异性研究和TREM2突变载体。 mol Neurodegener,2019,14:18-31 [18] Lanier LL,Corliss BC,Wu J等。 带有基于酪氨酸的活化基序的免疫受体DAP12参与激活NK细胞。 自然,1998,391:703-7 [19] Thornton P,Sevalle J,Deery MJ等。 trem2在H157-S158键上通过裂解脱落,以加速阿尔茨海默氏病相关的H157Y变体。 Embo Mol Med,2017,9:1366-78 [20] Piccio L,Buonsanti C,Cella M等。 识别J Immunol,2000,164:4991-5 [17] Del-Aguila JL,Benitez BA,Li Z等。TREM2脑转录本特异性研究和TREM2突变载体。mol Neurodegener,2019,14:18-31 [18] Lanier LL,Corliss BC,Wu J等。带有基于酪氨酸的活化基序的免疫受体DAP12参与激活NK细胞。自然,1998,391:703-7 [19] Thornton P,Sevalle J,Deery MJ等。trem2在H157-S158键上通过裂解脱落,以加速阿尔茨海默氏病相关的H157Y变体。Embo Mol Med,2017,9:1366-78 [20] Piccio L,Buonsanti C,Cella M等。识别


丙酸氟替卡松;沙美特罗辛萘甲酸酯
一般而言,FDA 的指导文件并不规定具有法律约束力的责任。相反,指导描述了该机构当前对某个主题的想法,除非引用特定的监管或法定要求,否则应仅将其视为建议。机构指导中的“应该”一词的使用意味着建议或推荐某事,但不是要求。活性成分:丙酸氟替卡松;沙美特罗昔萘酸酯剂型:粉末途径:吸入强度:0.1 mg/inh;EQ 0.05 mg 碱基/inh,0.25 mg/inh;EQ 0.05 mg 碱基/inh,0.5 mg/inh; EQ 0.05 mg 碱基/inh 推荐的研究:两种选择:(1) 四项体外生物等效性研究、一项比较特性研究和两项以药代动力学终点为目的的体内生物等效性研究,或 (2) 两项体外生物等效性研究、一项以药代动力学终点为目的的体内生物等效性研究和一项比较临床终点生物等效性研究 I. 选项 1:四项体外生物等效性研究、一项比较特性研究和两项以药代动力学终点为目的的体内生物等效性研究 为了通过此选项证明生物等效性,测试 (T) 产品与参考标准 (RS) 产品相比,在非活性成分或配方的其他方面应当没有差异,这些差异可能会显著影响活性成分的局部或全身利用度。例如,T 产品可以在质量 (Q1) 1 和定量 (Q2) 2 上与 RS 产品相同,以满足非活性成分没有差异。

1个upadacitinib(应用的新领域:克罗恩病,...
根据欧洲录取管理局(EMA)的要求,就制药企业家将要最小化的其他措施而言,培训材料是培训材料,这是医务人员和患者的信息(包括患者卡)提供。特别是培训和信息材料,其中包含有关upadacitib造成的可能副作用的说明,尤其是在包括TB和疱疹带状疱疹在内的严重和机会性感染以及出生错误(怀孕期间的风险),MACE,VTE,VTE和恶性疾病。

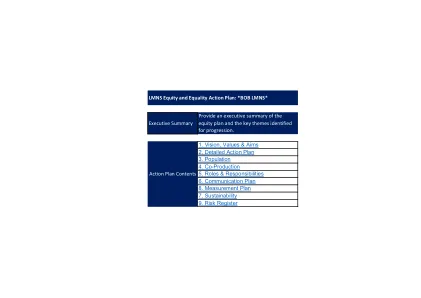

区分克罗恩病的新诊断策略...
背景:脂肪嫁接是重建武术中的高度用途,但具有不可预测的保留率和结果。与单独使用离心料的未经处理的脂肪酸或脂肪移植相比,该系统综述的主要概述是评估次级机械加工的脂肪酸是否有利地增强了脂肪移植物的血管生成潜力。次要结果是在比较上述组时评估围绕移植物的证据并改善结果。方法:在2022年2月之前,对MEDLINE,EMBASE和COCHRANE CENTRAL登记册进行了搜索。包括所有人类和动物研究,其中包括未加工,离心,次级机械碎片(SMF)或次级机械破坏(SMD)脂肪移植物之间的交叉比较。结果:包括31个全文。血管生成潜力。通过荧光激活的细胞分析(FACS)分析来定量间充质干细胞,血管血管干细胞和内皮祖细胞的细胞组成。脂肪移植的量保留率和有助于伤口愈合的脂肪移植率。尽管在某些研究的来源中,具有行业资助的研究的存在和方法学数据的报告不足,但数据显示,SMF移植物包含富集的周细胞种群,其血管腔内皮生长因子(VEGF)分泌增加。需要进一步的临床研究来评估人类研究中的潜在差异。动物研究表明,与离心移植物相比,SMD移植物可能会增加脂肪移植的率和伤口闭合率。但是,临床研究尚未显示出相似的结果。结论:在这项系统的综述中,我们能够得出结论,现有文献表明机械处理脂肪,无论是通过碎片或破坏,通过增强血管生长生长因子和相关的血管祖细胞水平来提高血管生成潜力。虽然体内动物研究稀缺,但综述的发现表明,次级机械脂肪会增强脂肪移植的保留率,并可以帮助伤口愈合。

CRISPR/Cas9基因编辑技术在阿尔茨海默病研究中的应用
[4] Kisilevsky R. 从关节炎到阿尔茨海默病:关于淀粉样变性发病机制的最新概念。Can J Physiol Pharmacol,1987,65:1805-15 [5] György B、Lööv C、Zaborowski MP 等人。CRISPR/Cas9 介导的瑞典 APP 等位基因破坏作为早发性阿尔茨海默病的治疗方法。Mol Ther Nucleic Acids,2018,11:429-40 [6] Zetterberg H、Mattsson N. 了解散发性阿尔茨海默病的病因。Expert Rev Neurother,2014,14:621-30 [7] Jack CR Jr、Knopman DS、Jagust WJ 等人。阿尔茨海默病病理级联动态生物标志物的假设模型。Lancet Neurol,2010,9:119-28 [8] Ittner LM、Ke YD、Delerue F 等。tau 的树突状功能介导阿尔茨海默病小鼠模型中的淀粉样蛋白 β 毒性。Cell,2010,142:387-97 [9] Muralidar S、Ambi SV、Sekaran S 等。tau 蛋白在阿尔茨海默病中的作用:主要的病理因素。Int J Biol Macromol,2020,163:1599-617 [10] Wang X、Wang W、Li L 等。阿尔茨海默病中的氧化应激和线粒体功能障碍。 Biochim Biophys Acta, 2014, 1842: 1240-7 [11] Grothe M, Heinsen H, Teipel SJ. 成年年龄范围内以及阿尔茨海默病早期阶段胆碱能基底前脑萎缩。Biol Psychiatry, 2012, 71: 805-13 [12] He Y, Ruganzu JB, Jin H, et al. LRP1 敲低通过调节 TLR4/NF- κB/MAPKs 信号通路加重 Aβ 1-42 刺激的小胶质细胞和星形胶质细胞神经炎症反应。Exp Cell Res, 2020, 394: 112166 [13] Huang HC, Hong L, Chang P, et al.壳寡糖减弱Cu 2+诱导的细胞氧化损伤和细胞凋亡,涉及Nrf2激活。Neurotox Res,2015,27:411-20 [14] Tomljenovic L. 铝和阿尔茨海默病:经过一个世纪的争论,是否存在合理的联系?J Alzheimers Dis,2011,23:567-98 [15] Shen H,Guan Q,Zhang X,等。阿尔茨海默病神经炎症的新机制:肠道菌群介导的NLRP3炎症小体的激活。Prog Neuropsychopharmacol Biol Psychiatry,2020,100:109884 [16] Ferreira-Vieira TH,Guimaraes IM,Silva FR,等。阿尔茨海默病:针对胆碱能系统。Curr Neuropharmacol,2016,14:101-15 [17] Scannevin RH。针对神经退行性蛋白质错误折叠障碍的治疗策略。Curr Opin Chem Biol,2018,44:66-74 [18] Giau VV,Lee H,Shim KH 等人。CRISPR-Cas9 的基因组编辑应用促进阿尔茨海默病的体外研究。Clin Interv Aging,2018,13:221-33 [19] Gupta D,Bhattacharjee O,Mandal D 等人。CRISPR-Cas9 系统:基因编辑的新曙光。生命科学, 2019, 232: 116636 [20] Makarova KS, Wolf YI, Alkhnbashi OS, et al.更新了