XiaoMi-AI文件搜索系统
World File Search System生物化学

CHEM-化学和生物化学
CHEM 105N 化学入门 (3 学分) 本课程是两个学期化学课程的第一部分,涵盖普通化学、有机化学和生物化学等主题。本部分将介绍无机(普通)化学的原理。涵盖的主题包括测量、原子和元素、化合物及其键、能量和物质、气体、溶液、酸和碱、化学反应和量、化学平衡和核化学。本课程不满足 CHEM 123N 的先决条件,不能用于 CHEM 专业或辅修。希望继续深造化学的学生应选修 CHEM 121N、CHEM 122N、CHEM 123N 和 CHEM 124N。如果学生已修过 CHEM 121N,则不允许修 CHEM 105N 的学分。CHEM 105N + CHEM 106N 满足大学科学本质通识教育要求的四个学分。先决条件:基本代数知识 共同要求:CHEM 106N

生物化学与分子生物学系
BCMB 430 - 分析生物化学和生物物理学 3 学分 课程目标:了解构成生物科学中使用的技术和仪器基础的物理科学原理 先决条件:生命科学学士学位课程。 第一单元 - 电化学技术和光度测定 11 小时 电化学的基本原理 - pH 电极 - 离子选择性 - 气体传感和氧电极 - 生物传感器的基本细节。比色法和分光光度法的原理和技术-比尔-朗伯定律-仪器-低色度和增色度-荧光测定-流式细胞术-原子吸收光谱法-圆二色性-光学旋光色散-核磁共振光谱-红外光谱第二单元-显微镜 7 小时显微镜-基本原理和应用-光-化合物-相衬-暗场-荧光显微镜扫描电子显微镜-透射电子显微镜 (TEM) -扫描隧道显微镜- (STM) -共聚焦显微镜。第三单元 - 离心 6 小时离心的基本原理 - 仪器、离心装置 - 离心机的类型 - 转子、配件 - 离心方法 - 沉降速度 - 沉降平衡 - 胶体 - 细胞分离方法。第四单元 – 色谱法 10 小时 色谱法的类型 - 柱色谱法、薄层色谱法、纸色谱法、吸附色谱法、分配色谱法、气液离子交换色谱法、亲和色谱法、高效液相色谱法 - 每种类型的原理 - 仪器和附件 - 检测方法和系统 - 定性和定量方面 - 应用;第五单元 – 电泳法 6 小时 电泳类型 - 纸和凝胶 - 琼脂糖和 PAGE - 脉冲场 - 毛细管 - 等电聚焦 - 印迹技术:西方、南方和北方。应用教科书 1. James, P. Allen. (2008). 生物物理化学,Wiley Blackwell,新泽西。2. Wilson, K. 和 Walker, J. (2010) 生物化学和分子生物学原理和技术,剑桥大学出版社,剑桥。推荐阅读 1. Horst, F. (2010) 基本一维和二维核磁共振波谱学,Wiley-VCH,新泽西。 2. Murphy, DB 和 Davidson, MW (2012) 光学显微镜和电子成像基础,Wiley-Blackwell,新泽西州。3. Freifelder, DM (1983) 物理生物化学 - 生物化学和分子生物学应用,WH Freeman,纽约


M.Phil的课程结果。生物化学
co 1:通过研究本文,学生将能够定义研究,解释和应用研究术语,描述研究过程以及与研究过程相关的主要活动,技能和道德。co 2:学生可以解释理论与研究之间的关系,描述和比较主要的定量和定性研究方法。CO 3:构建一项有效的研究建议,将作为研究项目的发射点,了解研究伦理的重要性,并将研究伦理纳入研究过程。CO 4:学生将轻松地使用统计工具和计算机软件来组织和分析数据。EBC 1:生物化学中的概念1:学生将获得有关不同代谢途径及其整合的知识。co 2:了解酶的性质以及酶 - 基底动力学的概念及其在生物反应中的重要性。CO 3:欣赏细胞成分,并将选择适当的基础生物学研究实验策略。CO 4:了解基本水平的免疫学及其生化重要性。EBC 2:人类生理学和临床生物化学CO 1:本课程将在人类生理学和临床生物化学方面提供合理的基础,以详细及其适当的水平深入了解所有身体系统的生理过程。co 2:学生将能够解释器官的活动是如何集成的,以提高效率。co 3:学生将准备确定正常生理学的变化如何导致疾病,并支持在健康和医学科学或相关领域的进一步研究。CO 4:本文还将提供对人体激素作用的理解,以调节不同器官系统的正常生理活性以及代谢过程。



化学和生物化学 - 本科课程
化学和生物化学系提供了四个学士学位的研究计划,并提供了两个学士学位和硕士学位。他们是化学学士学位,化学理学学士 - 美国化学学会认证,生物化学理学学士学位 - 美国化学学会认证,生物化学理学学士学位,是化学科学硕士学位 - 化学科学硕士学位 - 化学学士学位,具有生物化学科学学士学位,以及与科学硕士学位的生物学科学学士学位。

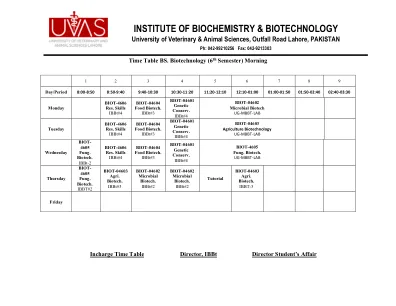
生物化学与生物技术研究所
日/周期8:00-8:50 8:50-9:40 9:40-10:30 10:30-11:20 11:20-12:10 12:10-01:00 01:00-01:50 01:50 01:50-02:40 02:40 02:40-02:40-03:30-03:30

生物化学教科书Stryer
杰里米·伯格(Jeremy M.他在1986年至1990年成为助理教授之前,在约翰·霍普金斯大学医学院与卡尔·帕博(Carl Pabo)完成了博士后奖学金。伯格随后搬到约翰·霍普金斯(Johns Hopkins),担任生物物理与生物物理化学系主教和主任,他一直担任该职位。后来,他成为美国国立卫生研究院国家一般医学科学研究所主任。2011年,伯格(Berg)加入了匹兹堡大学,担任计算与系统生物学教授和个性化医学研究所主任。在任职期间,他从2011年至2013年担任美国生物化学和分子生物学学会主席。Berg是美国科学发展协会的会员,也是美国国家科学院医学研究所的成员。他获得了几项著名的奖项,包括美国化学化学化学学会奖和Eli Lilly生物化学基础研究奖。此外,他还获得了来自各种组织的公共服务奖,并获得了许多教学奖。Berg还是Stephen J. Lippard的生物无机化学原理的合着者。斯坦福大学细胞生物学名誉教授Lubert Stryer教授在生物化学研究方面具有杰出的职业。斯坦福大学细胞生物学名誉教授Lubert Stryer教授在生物化学研究方面具有杰出的职业。他获得了医学博士学位来自哈佛医学院,并在克里斯托弗·沃尔什(Christopher T. Walsh)的监督下完成了博士后奖学金。Stryer目前是GlaxoSmithkline心力衰竭发现表现单元的高级科学研究员。著名的教授获得了众多奖项,包括2006年的国家科学勋章以及选举授予美国国家科学院和美国哲学学会。他对生物化学的贡献获得了Eli Lilly的生物化学基础研究奖和知识产权所有者协会的杰出发明人奖。Stryer关于生物化学的开创性工作一直是教育的基石,尤其是他于1975年出版的第一版,彻底改变了生物化学教学。第九版生物化学仍然忠于原始文本的完整性,具有清晰的写作,创新的图形以及尖端研究技术的覆盖范围。为了满足该主题的复杂性,第九版重点介绍了三个关键领域:整合文本和媒体,促进有效的问题解决方案,并提供积极学习的工具。包含Saplingplus数字解决方案为学生提供了一个独特的机会,可以通过交互式媒体丰富的资源来形象化复杂的生物化学概念。内置的评估,提示和有针对性的反馈使学生能够成为熟练的问题解决者。已引入新功能,以促进引人入胜的课堂环境。工具和资源现在可以在文本,Saplingplus和讲师材料中可用,使教师能够创建一个更活跃的学习空间。