XiaoMi-AI文件搜索系统
World File Search System甲基化

多囊卵巢综合症与表观遗传DNA甲基化调控的研究进展
多囊卵巢综合征(PCOS)是育龄妇女中最常见的内分泌疾病。尽管其发病率很高并且被认为是无排卵性不孕的主要原因,但人们对该综合征的了解仍然很少,仍存在诊断不足和治疗不足的情况,导致女性患者治疗方案的研究进展缓慢。这种复杂疾病的异质性是遗传、环境、内分泌和行为因素共同作用的结果。它通常与卵巢增大和功能障碍、雄激素水平升高和胰岛素抵抗有关。目前,尚无单一病因可以完全解释 PCOS 的发病机制。大多数证据表明 PCOS 是一种复杂的多因素疾病,具有高度的遗传性。表观遗传学是指基因组和基因表达的可遗传变化,而 DNA 序列没有任何改变。表观遗传学包括DNA甲基化、组蛋白修饰(乙酰化、磷酸化、甲基化等)和非编码RNA(ncRNA)含量的改变。现有研究认为表观遗传学,特别是DNA甲基化在PCOS的发病机制中起着至关重要的作用。

人类甲基化和非甲基化DNA集
产品描述人类甲基化和非甲基化的DNA集由两个对照DNA(非甲基化和甲基化)以及一组特定设计的引物,可与EZ DNA甲基化 - 甲基化 - 光点™结合使用,EZ DNA甲基化 - ez DNA甲基化基因甲基化基因甲基化基因,EZ DNA甲基化基因, Zymo研究以评估DNA的亚硫酸含量转化的效率。从包含DNA甲基转移酶DNMT1( - / - )和DNMT3B( - / - )1的细胞中纯化了人类HCT116 DKO非甲基化DNA。源自HCT116 DKO细胞的DNA具有低水平的DNA甲基化,可用作DNA甲基化分析的对照(图1)。人类HCT116 DKO甲基化的DNA被纯化为HCT116 DKO DNA,并且已在所有胞质位置进行了酶甲基化,该位置包括M.SSSI甲基转移酶2的CG二核苷酸,并可以用作DNA甲基化分析的阳性对照。在亚硫酸盐处理后,甲基化的胞嘧啶仍未转化(在哺乳动物中,胞嘧啶甲基化主要发生在CPG的情况下),而PCR后,非甲基化的胞嘧啶被转化为尿嘧啶并被检测为胸骨。DAPK1控制引物扩增了与死亡相关蛋白激酶1(DAPK1)基因的甲基化,非甲基化和混合甲基化拷贝,并用于在甲硫酸盐转化对照DNA后用于使用。恢复的DNA非常适合许多应用,包括下游分析,例如PCR,限制性核酸内切酶消化,测序等。
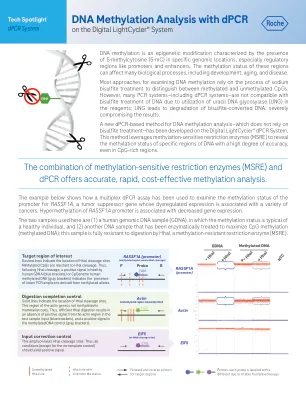
DNA甲基化分析DPCRDNA甲基化分析DPCR
高-GC区域(例如RASSF1A启动子)的扩增通常效率低下,即使在未甲基化的区域中也是如此(请参阅下图中图中的第一组数据;测试的GC的GC含量为72%GC)。这可能会使不论甲基化状态如何准确量化模板的数量。然而,在使用DigitalLightCycler®DPCR系统时,将高GC增强子添加到扩增反应中可以显着提高放大效率和定量准确性(请参见下文);这增加了基于DPCR的DNA甲基化分析的灵敏度和准确性。

1 甲基化与衰老的关系
表观遗传学涉及在不改变 DNA 序列的情况下调节遗传活动的分子修饰,这涉及多种细胞过程。衰老与损伤的积累有关,而损伤可能是由 DNA 甲基化引起的,DNA 甲基化是通过“表观遗传时钟”来标记生物年龄的。然而,文献仍然缺乏将表观遗传时钟、甲基化及其在衰老中的影响的主要方面联系起来的清晰的综合。因此,本研究旨在探讨 DNA 甲基化如何影响衰老,并系统化有关其与细胞代谢和健康衰老的关系的知识。使用的数据库是 PubMed,使用过滤器和排除标准后搜索选择了四篇文章。 DNA甲基化是表观遗传时钟的基础之一,因为它影响导致衰老的几种细胞机制。这些发现强调了将表观遗传学视为衰老和相关过程的重要因素的重要性。关键词:衰老。表观遗传学。甲基化。表观遗传时钟。知识领域:分子生物学。介绍


DNA甲基化与HIV
1心脏肺创新中心,不列颠哥伦比亚大学,温哥华,不列颠哥伦比亚省,加拿大2分子医学与治疗中心,不列颠哥伦比亚大学,加拿大不列颠哥伦比亚省温哥华大学,加拿大温哥华大学,3月3日,伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伊利诺伊州伦敦市的伊利诺伊州伊利诺伊州大学,伦敦大学,伦敦大学,Uniziz Uniziz Unceminiz Inter,Uniziz Inuce,Uniziz Intim,Uniziz in U. Poliklinik II,德国伍兹堡大学,沃兹堡大学6号柯比研究所,UNSW悉尼,悉尼,悉尼,新南威尔士州,新南威尔士州,澳大利亚7明尼阿波利斯退伍军人事务卫生保健系统,肺部,重症监护和睡眠医学和睡眠医学和肺部,急诊医学和校友,分子,分子,分子,分校,肺病,重症监护和睡眠医学,疾病和睡眠医学,


RNA甲基化,CRISPR-CAS和BEYER
莱斯利获得了A.B.哈佛大学的生物学博士学位,作为约翰·哈佛学者和A.M.和博士普林斯顿大学(Laura Landweber)的生物学博士学位和汤姆·缪尔(Tom Muir)作为佩特里(Petrie)的研究员,在那里他使用基因组学和生化分级方法来识别与RNA M6A甲基转移酶同源的新型DNA甲基转移酶配合物,Mettl3-14(Beh等,Cell et al。,Cell 2019)。 莱斯利随后在哥伦比亚大学的山姆·斯特恩伯格(Sam Sternberg)进行了简短的博士后工作,在那里他使用基因组,结构和生物化学方法研究CRISPR-CAS系统,介导RNA指导的DNA整合(Hoffmann*,Hoffmann*,Hoffmann*,hoffmann*,Kim*,Kim*,kim*et al。 之后,莱斯利(Leslie)从事行业职业,加入Illumina,领导一个研究小组开发新型表观遗传学测定的研究小组。 在进行生物学研究并对学生产生影响的愿望的驱动下,莱斯利(Leslie)于2022年9月回到*Star / imcb担任首席研究员。哈佛大学的生物学博士学位,作为约翰·哈佛学者和A.M.和博士普林斯顿大学(Laura Landweber)的生物学博士学位和汤姆·缪尔(Tom Muir)作为佩特里(Petrie)的研究员,在那里他使用基因组学和生化分级方法来识别与RNA M6A甲基转移酶同源的新型DNA甲基转移酶配合物,Mettl3-14(Beh等,Cell et al。,Cell 2019)。莱斯利随后在哥伦比亚大学的山姆·斯特恩伯格(Sam Sternberg)进行了简短的博士后工作,在那里他使用基因组,结构和生物化学方法研究CRISPR-CAS系统,介导RNA指导的DNA整合(Hoffmann*,Hoffmann*,Hoffmann*,hoffmann*,Kim*,Kim*,kim*et al。之后,莱斯利(Leslie)从事行业职业,加入Illumina,领导一个研究小组开发新型表观遗传学测定的研究小组。在进行生物学研究并对学生产生影响的愿望的驱动下,莱斯利(Leslie)于2022年9月回到*Star / imcb担任首席研究员。

甲基化操作向导(Meow)
图1。Meow在长阅读测序数据中识别差异甲基化区域。A. Meow需要一组带有填充的MM和ML标签的对齐的BAM文件以及包含感兴趣区域列表的床文件,例如CPG岛,以构建参考数据库。在构建参考数据库后,可以在参考队列中执行一项输出分析,以识别该数据集中的唯一差异甲基化区域(DMR)。也可以使用已经构建的参考数据库来识别DMR的测试样本运行。两种方法的输出都在表或图形格式中获得。B.与已知具有Prader-Willi综合征的测试样品相比,与19个随机样品的对照数据库相比,显示了已知具有Prader-Willi综合征的测试样品的显着差异甲基化的位点(红色),该数据库是1000个基因组项目ONT测序联盟的一部分。C. Meow生成图形,说明了测试样品和对照数据集之间甲基化频率的显着差异。所示的五个DMR表示(b)中的显着值。D.色带图显示了查询中每个C和G的甲基化频率,相对于控制数据库甲基化频率在同一位置的平均值和标准误差。

低温SABATIER CO2甲基化
Brodie于1872年首先描述了CO/CO 2与H 2的混合物与H 2的混合物。[1]三十年后,1902年,“法国正世俗主义”的促进者保罗·萨巴蒂尔(Paul Sabatier)和父亲让·巴蒂斯特·森德伦斯(Jean Baptiste Senderens)[2]描述了他们与CO和CO 2氢化对镍催化剂的反应有关的发现。[3]两种流室MIC反应有选择性地产生甲烷。在镍上,反应在250°C下轻松进行,而在CO 2的情况下需要350°C。[4]使用较高的温度引起的碳沉积。使用钴的使用暗示在较高温度下起作用,以开始反应。几年后,萨巴蒂尔(Sabatier)以有远见的方式提议将这种反应应用于CO 2和电解产生的氢气的产生或加热气体。[5]部分要归功于这些关于CO 2甲基化的研究(今天也称为Sabatier反应),Sabatier于1912年与Victor Grignard一起获得了化学诺贝尔奖。从历史上看