XiaoMi-AI文件搜索系统
World File Search System磷酸



磷酸锂电池的回收
o专用拆卸和预处理(准确)o电解质材料(ENEA)o黑色质量(fraunhofer ilt)的在线表征o恢复石墨(SIM2和VITO)o氢化透明液含量锂的液化液和转换型液化液材料材料的液化材料(Simdode)的液化材料(Simdody ligrade lith infirnim infirate ligrade ligrane ligrate pirever infre firgin hydohydrate otect ofirn hydrox preight pirever pireven of inf tigner of inf pirect of。 (vito)

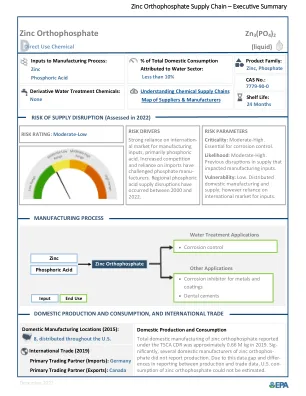
锌磷酸锌供应链
在2019年,在CDR下报告的锌高磷酸锌的全部制造约为0.66 m kg(EPA,2020年);但是,一些国内制造商声称拥有机密的业务信息,并且没有向EPA报告生产量,其中包括历史悠久的重要国内制造商ICL特色产品和撒切尔公司。大多数锌邻磷酸锌生产设施生产用于金属涂料的锌邻磷酸锌混合物。Carus Corporation是2019年水处理报告生产的最大的高磷酸锌混合物。图2中所示的国内制造地点数量代表了2015年的运营设施(EPA,2016年)。NSF/ANSI标准60认证的量磷酸锌供应用于饮用水处理的锌在美国广泛分布(NSF International,2021年)。有关制造地点和供应商地点的最新清单,请访问美国环境保护局(EPA)的化学定位器工具(EPA,2022a)。

磷酸铁锂电池用户操作手册
9 安装 ................................................................................................................................................ 18

磷酸铁锂(LFP)48MPLhE100-16S
• 系统监控电池和模块的电压、电流和温度。内置放电和充电过流、过热、低温、低压和高压以及短路保护 • 通过 RS485 和 Modbus 进行 BMS 维护和服务通信,可与逆变器和其他设备轻松连接 • 通过干触点实现 2 级远程报警

牙本质唾液磷酸蛋白信号...
牙本质生成始于成牙本质细胞,成牙本质细胞合成并分泌非胶原蛋白 (NCP) 和胶原蛋白。当牙本质受伤时,牙髓祖细胞/间充质干细胞 (MSC) 可以迁移到受伤区域,分化为成牙本质细胞并促进反应性牙本质的形成。牙髓祖细胞/MSC 分化在给定的生态位中受到控制。在牙齿 NCP 中,牙本质唾液酸磷蛋白 (DSPP) 是小整合素结合配体 N 连接糖蛋白 (SIBLING) 家族的成员,该家族的成员具有共同的生化特征,例如 Arg-Gly-Asp (RGD) 基序。DSPP 表达具有细胞和组织特异性,在成牙本质细胞和牙本质中高度常见。DSPP 突变会导致遗传性牙本质疾病。 DSPP 在蛋白水解作用下被催化成牙本质糖蛋白 (DGP)/唾液酸蛋白 (DSP) 和磷蛋白 (DPP)。DSP 进一步加工成活性分子。DPP 包含 RGD 基序和丰富的 Ser-Asp/Asp-Ser 重复区。DPP-RGD 基序与整合素 αVβ3 结合,并通过丝裂原活化蛋白激酶 (MAPK) 和粘着斑激酶 (FAK)-ERK 通路激活细胞内信号传导。与其他 SIBLING 蛋白不同,DPP 在某些物种中缺乏 RGD 基序。然而,DPP Ser-Asp/Asp-Ser 重复区与磷酸钙沉积物结合,并通过钙调蛋白依赖性蛋白激酶 II (CaMKII) 级联促进羟基磷灰石晶体生长和矿化。DSP 缺乏 RGD 位点,但含有信号肽。信号域的三肽与内质网内的货物受体相互作用,促进 DSPP 从内质网运输到细胞外基质。此外,DSP 的中间和 COOH 末端区域与细胞膜受体、整合素 β6 和闭合蛋白结合,诱导细胞分化。本综述可能揭示 DSPP 在牙发生过程中的作用。

石墨/铁磷酸锂-ARPI
锂离子电池的准确建模对于从电动汽车(EV)到网格存储的一系列AP平板优化性能和安全至关重要。本文使用60 AH Prismatic石墨/锂磷酸铁电池作为案例研究,对两种普遍的电池建模方法进行了两种普遍的电池建模方法:等效电路模型(ECM)和基于物理的模型(PBM)。这项工作的重点是通过在恒定和可变的电流密度下的不同环境温度下的一组全面的电气测试(包括全球协调的轻型车辆测试周期(WLTC)协议),通过在不同环境温度下进行全面的电气测试来开发,参数化和交叉验证这些方法。此评估不仅评估了ECM和PBM的准确性和可靠性,还强调了其优势和局限性。ECM在其校准范围内和可变电流轮廓内显示了计算速度,易于校准和准确性的优势。然而,其准确性在较高的电流下会降低,尤其是对于延长的电流脉冲以及校准范围之外的延长,这在1C以上的充电方案中证明了这一点。相反,PBM在校准数据集之外保持准确性,但需要估计许多物理参数,艰苦的校准过程以及用于可变当前情况的扩展计算时间。在所研究的条件范围内(从C/3到2C之间的10℃和40℃),ECM的电压预测的平均误差为51.5 mV,PBM的平均误差为19.3 mV,而ECM的平均误差为0.9℃,而对于温度预测,PBM的平均值为0.9°C。总而言之,虽然ECM适用于以短暂和低强度的电荷脉冲来重现恒定放电或类似WLTC的轮廓,但PBM强度在于其对高速运营的预测性,使其成为模拟现实的EV负载操作和优化快速收费协议的互补工具。这些见解有助于电池技术的持续发展,重点是现实且适用的模型开发和参数化。
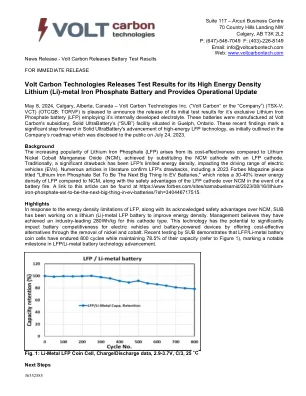
金属磷酸铁电池并提供
背景是磷酸锂(LFP)的普及,与锂镍钴锰氧化物(NCM)相比,其成本效益引起,通过用LFP阴极代替NCM阴极来实现。传统上,LFP的能量密度有限,影响了电动汽车(EV)的驱动范围。文献中的许多文章证实了LFP的缺点,包括2023年《福布斯》杂志的文章,标题为“磷酸锂,将是电动电动电池中的下一件大事”,它指出,与NCM相比,LFP的LFP能量密度降低了30-40%,与NCM相比,LFP天主教徒与NCM的安全优势相比。A link to this article can be found at https://www.forbes.com/sites/samabuelsamid/2023/08/16/lithium- iron-phosphate-set-to-be-the-next-big-thing-in-ev-batteries/?sh=340446717515.