XiaoMi-AI文件搜索系统
World File Search System等位基因
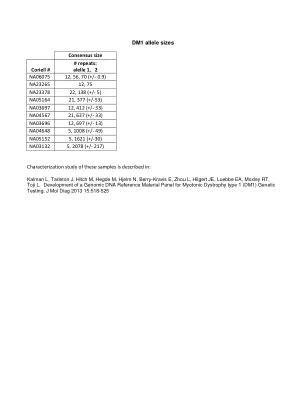
DM1 等位基因大小
Coriell # # 重复:等位基因 1、2 NA06075 12、56、70 (+/‐ 0.9) NA23265 12、75 NA23378 22、138 (+/‐ 5) NA05164 21、377 (+/‐53) NA03697 12、412 (+/‐ 33) NA04567 21、637 (+/‐ 33) NA03696 12、697 (+/‐ 13) NA04648 5、1008 (+/‐ 49) NA05152 5、1621 (+/‐30) NA03132 5、2078 (+/‐ 217)

使用等位基因特异性PCR
这项研究范围内的改进是可能的,包括将测定法应用于早期繁殖线,最初通过病原体测试验证标记呼叫和更快的提取技术。KASP标记如果我们可以在人口达到近亲状态之前为其选择它们,将特别有用。f2至F4选择是理想的选择,因为杂合子和那些纯合子与易感性连接标记等位基因的纯合子可以立即被丢弃。在随后的几代人中较少的植物/线条将节省繁殖计划中的时间和空间。病原体测试

CYP2D7 混合等位基因分型
CYP2D6 是一种非常重要的药物基因,因为它负责 20% 至 30% 临床使用药物的代谢或生物活化。然而,尽管它的长度相对较短(只有 4.4 kb),但由于与邻近假基因的高度相似性以及 CYP2D6-CYP2D7 杂交的频繁出现,它是基因分型最困难的药物基因之一。不幸的是,大多数当前的基因分型方法无法正确确定完整的 CYP2D6-CYP2D7 序列。因此,我们开发了一种基因分型检测方法,通过优化无 PCR 纳米孔 Cas9 靶向测序 (nCATS) 方法与自适应测序相结合,并开发了一种新的综合长读基因分型 (CoLoRGen) 流程,以生成复杂区域的完整等位基因特异性共识序列。 CoLoRGen 流程首先生成两个等位基因的一致序列,然后确定大结构变异和小变异,最终分配正确的星号等位基因。在参考样本中,我们的基因分型检测证实了 CYP2D6-CYP2D7 大结构变异、单核苷酸变异 (SNV) 以及小插入和缺失 (INDEL) 的存在,而这些是大多数当前检测无法检测到的。此外,我们的结果提供了直接证据,表明 NA12878 DNA 的 CYP2D6 基因型应更新为包括 CYP2D6-CYP2D7 * 68 杂交和与现有参考相比的几个额外的单核苷酸变异。最终,nCATS-CoLoRGen 基因分型检测还可以通过检测和分期从头突变以及已知的大结构变异和小变异,从而实现更准确的基因功能预测。
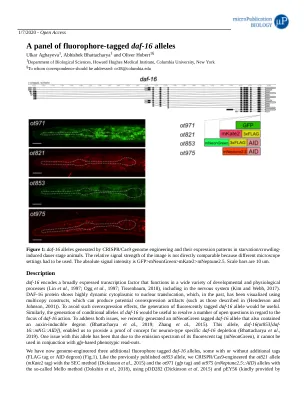
一组荧光标记的 daf-16 等位基因
daf-16 编码一种广泛表达的转录因子,在多种发育和生理过程中发挥作用 (Lin et al., 1997; Ogg et al. , 1997; Tissenbaum, 2018),包括在神经系统中 (Kim and Webb, 2017)。DAF-16 蛋白表现出高度动态的细胞质到核易位,过去曾使用多拷贝构建体进行可视化,这可能会产生潜在的过表达伪影(例如 (Henderson and Johnson, 2001) 中描述的那些)。为了避免这种过表达效应,生成荧光标记的 daf-16 等位基因将很有用。同样,生成 daf-16 的条件等位基因将有助于解决有关 daf-16 作用重点的许多悬而未决的问题。为了解决这两个问题,我们最近生成了一个带有 mNeonGreen 标记的 daf-16 等位基因,该等位基因还包含一个生长素诱导的降解子 (Bhattacharya 等人,2019;Zhang 等人,2015)。该等位基因 daf-16(ot853[daf- 16::mNG::AID]) 使我们能够为神经元类型特异性 daf-16 耗竭提供概念验证 (Bhattacharya 等人,2019)。该等位基因的一个问题是,由于其荧光标记 (mNeonGreen) 的发射光谱,它不能与基于 gfp 的表型读数结合使用。

高质量基因组组装使得能够预测等位基因...
1 北京林业大学生物科学与技术学院, 国家林木育种与生态修复工程研究中心, 林木分子设计育种北京市高精尖创新中心, 林木育种国家工程实验室, 林木园林植物遗传育种教育部重点实验室, 北京 100083 2 山东省农业科学院作物种质资源研究所, 作物遗传改良与生理生态重点实验室, 济南 250100 3 广东省农业科学院水稻研究所, 农业农村部南方优质水稻遗传育种重点实验室 (省部共建) , 广东省水稻育种新技术重点实验室, 广州 510640 4 宁夏大学农学院, 银川 750021 5 云南省水稻遗传改良重点实验室中国科学院昆明植物研究所东亚植物多样性与生物地理学重点实验室极小种群植物综合保护重点实验室,云南昆明 650201 6 山东农业大学林学院,山东泰安 271000 7 于默奥大学生态与环境科学系于默奥植物科学中心,瑞典于默奥 SE-901 87 8 不列颠哥伦比亚大学林业与保护科学系,加拿大不列颠哥伦比亚省温哥华,V6T 1Z4 9 图能森林遗传研究所,德国格罗斯汉斯多夫 22927 10 根特大学植物生物技术和生物信息学系,比利时根特 9052 11 VIB 植物系统生物学中心,比利时根特 9052 12 微生物生态学和基因组学中心,比勒陀利亚大学生物化学、遗传学和微生物学系,比勒陀利亚 0028,南非 13 南京农业大学园艺学院,高等交叉研究院,南京 210095,中国 14 于默奥植物科学中心,植物生理学系,于默奥大学,SE-901 87 于默奥,瑞典 15 森林与森林科学系,Faculté de林业,地理与地理,拉瓦尔大学,魁北克,QC G1V 0A6,加拿大

来自外来种质的强效等位基因的良好例子
• 80% 的玉米基因组被打碎了,重复的逆转录病毒序列 • 去除重复序列后,数千万个单核苷酸多态性 • 广泛的结构变异(一个品种与另一个品种相比,缺少大量 DNA) • 一些性状(例如,种子颜色)由影响巨大的单个序列变异控制 • 大多数性状由数十到数千个序列变异控制,并与环境有复杂的相互作用

跨等位基因频谱的人类代谢基因图谱
。CC-BY-NC-ND 4.0 国际许可证 它是永久可用的。 是作者/资助者,已授予 medRxiv 许可以显示预印本(未经同行评审认证)预印本 此版本的版权所有者于 2025 年 2 月 2 日发布。;https://doi.org/10.1101/2025.01.30.25321073 doi:medRxiv 预印本

高质量基因组组装使得能够预测等位基因...
1 北京林业大学生物科学与技术学院, 国家林木育种与生态修复工程研究中心, 林木分子设计育种北京市高精尖创新中心, 林木育种国家工程实验室, 林木园林植物遗传育种教育部重点实验室, 北京 100083 2 山东省农业科学院作物种质资源研究所, 作物遗传改良与生理生态重点实验室, 济南 250100 3 广东省农业科学院水稻研究所, 农业农村部南方优质水稻遗传育种重点实验室 (省部共建) , 广东省水稻育种新技术重点实验室, 广州 510640 4 宁夏大学农学院, 银川 750021 5 云南省水稻遗传改良重点实验室中国科学院昆明植物研究所东亚植物多样性与生物地理学重点实验室极小种群植物综合保护重点实验室,云南昆明 650201 6 山东农业大学林学院,山东泰安 271000 7 于默奥大学生态与环境科学系于默奥植物科学中心,瑞典于默奥 SE-901 87 8 不列颠哥伦比亚大学林业与保护科学系,加拿大不列颠哥伦比亚省温哥华,V6T 1Z4 9 图能森林遗传研究所,德国格罗斯汉斯多夫 22927 10 根特大学植物生物技术和生物信息学系,比利时根特 9052 11 VIB 植物系统生物学中心,比利时根特 9052 12 微生物生态学和基因组学中心,比勒陀利亚大学生物化学、遗传学和微生物学系,比勒陀利亚 0028,南非 13 南京农业大学园艺学院,高等交叉研究院,南京 210095,中国 14 于默奥植物科学中心,植物生理学系,于默奥大学,SE-901 87 于默奥,瑞典 15 森林与森林科学系,Faculté de林业,地理与地理,拉瓦尔大学,魁北克,QC G1V 0A6,加拿大

定义适合等位基因选择性治疗的患者...
针对体细胞旁观者遗传事件的疗法代表了癌症治疗的新途径。我们最近发现了一组结直肠癌 (CRC) 患者,他们对一个野生型和一个低活性等位基因 (NAT2*6) 是杂合的,但由于 8p22 的杂合性缺失 (LOH),他们的肿瘤中缺少野生型等位基因。这些肿瘤对用 NAT2 的细胞毒性底物(6-(4-氨基苯基)-N-(3,4,5-三甲氧基苯基)吡嗪-2-胺,APA)治疗敏感,并指出 NAT2 缺失是 CRC 肿瘤在治疗上可利用的弱点。为了更好地估计可治疗的 CRC 患者的总数,我们在此确定了 LOH 后还保留其他 NAT2 低活性变体的肿瘤细胞是否对 APA 治疗有反应。发现普遍存在的低活性等位基因 NAT2*5 和 NAT2*14(而非 NAT2*7)是低代谢物,对 APA 具有高敏感性。通过分析两个不同的 CRC 患者队列,我们在约 24% 的肿瘤中检测到 APA 可靶向的 NAT2 等位基因的杂合性以及指向 LOH 的等位基因失衡。最后,为了在临床环境中对肿瘤和患者匹配的正常样本中的 NAT2 基因座进行单倍型分析,我们开发并展示了一种基于长读测序的检测方法。每年共有 > 79,000 名 CRC 患者符合对 NAT2 LOH 疗法具有高敏感性的遗传标准,并且可以通过临床测序评估他们的资格。

CAS9生成条件小鼠等位基因的方法
1。我们对Yang等人发表的MECP2基因座的结果。已通过Jaenisch(8 - 10%正确的等位基因),Yang(8%正确的等位基因)和Hatada的组(2 - 6%正确等位基因)[3]的独立实验复制。此外,多个同行评审的出版物[3-7]成功使用了此方法来创建条件敲除(CKO)小鼠(在11个基因座中有9个成功,效率为2.5%至18%)。我们注意到,CRISPR/ CAS9生成CKO小鼠的效率可能会有所不同,这可能是由于平台特征或实验条件的不同。2。Gurumurthy等人使用的条件。[1]与我们论文中使用的条件不符。Gurumurthy等人使用的CRISPR试剂的浓度。 '在MECP2基因座上的研究[1](Cas9 mRNA的10 ng/μL,SGRNA的10 ng/μL,寡核素的10 ng/μL)比Yang等人所用的 sgrNA的RNA和10 ng/μL)。 ' s实验(CAS9 100 ng/μL,SGRNA 50 ng/μL和100 ng/μL的实验)[2]和Yang等。 ' s先前[8]和以下出版物[9-12]。 众所周知,CRISPR试剂的浓度与基因组编辑效率密切相关。 3。 我们在原始论文中使用了压电驱动的合子注入方法,该方法允许以更高的浓度注入CRISPR组件。 Gurumurthy等人使用的该方法和前核注射方法之间的差异。 也可能有助于成功的利率差异。sgrNA的RNA和10 ng/μL)。 's实验(CAS9 100 ng/μL,SGRNA 50 ng/μL和100 ng/μL的实验)[2]和Yang等。 's先前[8]和以下出版物[9-12]。众所周知,CRISPR试剂的浓度与基因组编辑效率密切相关。3。我们在原始论文中使用了压电驱动的合子注入方法,该方法允许以更高的浓度注入CRISPR组件。Gurumurthy等人使用的该方法和前核注射方法之间的差异。也可能有助于成功的利率差异。