XiaoMi-AI文件搜索系统
World File Search System纺锤体

低剂量药物组合预防纺锤体极...
摘要 有丝分裂在基于微管的纺锤体控制下,是抗癌治疗的一个有吸引力的靶点,因为癌细胞会经历频繁且不受控制的细胞分裂。破坏有丝分裂的微管靶向剂或有丝分裂激酶或微管马达的单分子抑制剂可以高效杀死癌细胞。然而,这些治疗方法存在严重的缺点:它们还针对经常分裂的健康组织,例如造血系统,并且由于原发性或获得性耐药机制,它们经常失去效力。在癌细胞分裂中出现的另一个目标是它们将有丝分裂纺锤体的极点“聚集”成双极结构的能力。这种机制对于癌细胞的特定存活是必要的,这些癌细胞由于经常存在异常的着丝粒数目或其他纺锤体缺陷而倾向于形成多极纺锤体。在这里,我们讨论了针对纺锤体极点聚集的组合治疗的最新发展,这些治疗专门针对具有异常着丝粒数目的癌细胞,并且由于其组合性质,有可能避免耐药机制。

特定染色体的非整倍性对缺乏纺锤体检查点蛋白BUB3
急性呼吸道感染(ARIS)是整个生命周期发病率和死亡率的主要原因。在全球范围内,估计每年发生的170亿ARI,占儿童死亡人数240万人(> 740,000人死亡)[1]。尽管这些ARI中的大多数是由呼吸道病毒引起的,但大多数严重或致命的病例是由细菌呼吸道病原体引起的。越来越多地,上呼吸道(URT)中的共生微生物会影响呼吸道病毒感染的风险和严重程度,以及细菌病原体对定殖和感染的抗性。因此,人们对利用这些微生物 - 微生物或微生物 - 主机相互作用的兴趣越来越多,以制定新的ARI策略或治疗[2]。尽管益生菌的现代历史可以追溯到一个多世纪以来,但活细菌菌株的鼻内给药将代表我们预防和治疗ARIS的方法的转变。这种鼻腔益生菌的必要特征将包括粘附上皮并成功地定居人类的能力,缺乏对呼吸性上皮细胞的细胞毒性,对地平线基因转移和移动遗传元件的某种程度的抵抗力,低倾向,低倾向,可侵犯宿主组织,使宿主的组织以及可用的可用抗药性可用的抗生素。下面,我们描述了一种研究的细菌物种,即dolosigranulum pigrum,越来越多地将其视为人类URT中的基石物种,也被视为预防ARI预防或治疗的有希望的鼻益生菌候选者。

有丝分裂纺锤体中协同微管和动粒动力学的双稳态和振荡
摘要。在有丝分裂纺锤体中,微管在中期通过捕获键附着在动粒上,微管解聚力引起随机染色体振荡。我们研究了纺锤体模型中的协同随机微管动力学,该模型由一组平行微管组成,这些微管通过弹性接头附着在动粒上。我们包括微管的动态不稳定性以及弹性接头对微管和动粒的作用力。采用基于福克-普朗克方程的平均场方法,对外力作用于动粒的单侧模型进行分析求解。该解建立了微管集合的双稳态力-速度关系,与随机模拟一致。我们推导出双稳态的接头刚度和微管数的约束。单侧纺锤体模型的双稳态力-速度关系导致双侧模型中的振荡,这可以解释中期随机染色体振荡(方向不稳定性)。我们推导出中期染色体振荡的连接体刚度和微管数的约束。将极向微管通量纳入模型,我们可以解释实验观察到的极向通量速度高的细胞中染色体振荡的抑制。然而,在存在极向喷射力的情况下,染色体振荡持续存在,但幅度减小,姊妹动粒之间有相移。此外,极向喷射力是必要的,以使染色体在纺锤体赤道处对齐,并稳定两个动粒的交替振荡模式。最后,我们修改了模型,使得微管只能对动粒施加拉力,从而导致两个微管集合之间发生拉锯战。然后,到达动粒后诱发的微管灾难是刺激振荡的必要条件。该模型可以定量再现 PtK1 细胞中动粒振荡的实验结果。

在 PRC1 和 EB1 基因编辑细胞中,中央纺锤体的逐渐压缩降低了其动态性
在有丝分裂过程中,纺锤体会发生形态和动态变化。它在后期开始时重组,此时反平行束 PRC1 积累并将中央纺锤体蛋白募集到中间区。人们对中央纺锤体在人类细胞中形态变化过程中的动态特性如何变化知之甚少。利用基因编辑,我们生成了从其内源性荧光位点表达 PRC1 和 EB1 的人类细胞,以量化其天然纺锤体分布和结合/解离周转。EB1 正末端追踪显示微管生长普遍减慢,而 PRC1 与其酵母直系同源物 Ase1 类似,与压缩的反平行微管重叠结合越来越强。 KIF4A 和 CLASP1 与中央纺锤体的结合更具动态性,但也显示出减慢的周转速度。这些结果表明,中央纺锤体在有丝分裂过程中逐渐变得更加稳定,这与最近在有丝分裂后期中央纺锤体中反向平行中区束形成的“捆绑、滑动和压缩”模型一致。

PICH 通过其 DNA 转位酶和 SUMO 相互作用活动影响纺锤体组装检查点
致作者的评论(必填):在本稿中,Lama 及其同事认为 PICH 重塑了 SUMO 化蛋白,以确保纺锤体组装检查点的正确暂时沉默。支持这一想法的主要观察结果是,PICH 的消耗,或在缺乏内源性 PICH 的细胞中重新表达缺乏 SUMO 结合能力或 ATPase 活性的外源性 PICH 突变体(分别被识别为 PICH ∆3SIM 和 K128A)在有丝分裂中(非常轻微地)延迟。作者询问这种短暂的停滞是否是由 Topo2alpha 依赖性通路的激活引起的(在之前的论文中进行了描述,并命名为 TRC,代表 Topo2alpha 响应检查点)。在得出事实并非如此的结论后,他们转向纺锤体组装检查点 (SAC),并发现在 PICH 消耗时或在表达功能失调的 PICH 突变体的细胞中,检查点蛋白 MAD1 在动粒上的停留时间延长。由于已知 PICH 会与 SUMO 化蛋白相互作用,作者推测 PICH 的缺失或用突变体替代可能导致 SUMO 化蛋白的积累,这可能是观察到的有丝分裂延迟的原因。为了验证这个想法,作者生成了一个表达标记 SUMO2 的细胞系,并比较了在存在或不存在 PICH 功能的情况下 SUMO2 结合蛋白的丰度。这确定了几种蛋白质,当 PICH 功能受损时,它们的 SUMO 化似乎会增加。在这些蛋白质中,作者确定了 BUB1,并证明在 PICH 缺失后 BUB1 动粒水平略有增加,这种影响可能是由于检查点激活恢复缺陷造成的。作者的模型是 PICH 有助于从动粒中去除 SUMO 化蛋白以促进检查点沉默。本文介绍的工作是通过创建几个细胞系实现的,清楚地反映了作者的大量宝贵努力。这项研究的主要局限性在于,观察到的影响非常小,并且没有最终证据表明导致这些影响的 PICH 的功能是精确且完全调节性的。它可能反映出持续的小附着错误,可能是由着丝粒染色质组织中的小问题引起的,该问题会向 SAC 发出信号。也就是说,延迟可能不只是反映出沉默错误,而是持续的检查点激活,这是作者没有解决的问题,而且考虑到停滞的实体很小,这个问题很难解决。在这方面,提出的模型也将过度的 SUMO 化确定为有丝分裂延迟的原因,虽然并非难以置信,但在分析的这个阶段似乎没有得到充分支持。在没有 PICH 的情况下观察到 SUMO 化增加,但细胞能够在对照细胞之后几分钟离开有丝分裂,这意味着必须存在处理过量 SUMO 的其他蛋白质。由于作者没有排除有丝分裂延迟仅仅是由真正的 SAC 激活引起的,PICH 在控制 SUMO 化方面的作用仍不确定。因此,总的来说,我认为这项研究虽然很有价值,但尚未代表完全令人信服的概念或机制进步。其他问题 - 图 1c 和 2c 中 ∆PICH 细胞中有丝分裂时间的差异引发了一致性问题。为什么这两种情况下有丝分裂退出的时间不同? - 在图 3 中,∆PICH 细胞中动粒处 MAD1 的持续时间远远超过 50 分钟,即远远超过这些细胞退出有丝分裂所需的时间(约 35 分钟,如图 1 所示)。这似乎相当难以置信,因为 MAD1 从动粒处的丢失总是先于有丝分裂退出。次要观点 -图 1B:最后一行,第 5 个面板,右下角部分隐藏的文本 -图 1C:如果作者指出此图中所示各种条件下有丝分裂退出的平均时间,将会很有帮助。 -在文本和相关图中指出 TopoIIalpha 带有 FLAG 标记

串行部分电子断层扫描和3D重构有丝分裂纺锤体中微管组织的定量分析
为了分析有丝分裂过程中细胞结构的分析,需要纳米分辨率来可视化纺锤体中微管的组织。在这里,我们提出了一种详细的方案,可用于在培养物中生长的细胞中整个有丝分裂纺锤体的3D重建。为此,我们将富含有丝分裂阶段的哺乳动物细胞附着在蓝宝石盘上。我们的协议进一步涉及通过高压冻结,冻结固定和树脂嵌入的冷冻污染。然后,我们使用荧光光学显微镜在树脂包裹的样品中选择有丝分裂细胞。接下来是大规模电子断层扫描,以重建3D中所选的有丝分裂纺锤体。然后,生成和缝合的电子断层图用于半自动分段微管,以进行纺锤体组织的随后定量分析。因此,通过提供详细的相关光和电子显微镜(CLEM)方法,我们为细胞生物学家提供了一种工具集来简化纺锤体微管的3D可视化和分析(http://kiewisz.shinyapps.io/asga)。此外,我们指的是一个最近启动的平台,该平台允许交互式显示3D重建有丝分裂纺锤体(https://cfci.shinyapps.io/asga_3dviewer/)。

串行部分电子断层扫描和3D重构有丝分裂纺锤体中微管组织的定量分析
为了分析有丝分裂过程中细胞结构的分析,需要纳米分辨率来可视化纺锤体中微管的组织。在这里,我们提出了一种详细的方案,可用于在培养物中生长的细胞中整个有丝分裂纺锤体的3D重建。为此,我们将富含有丝分裂阶段的哺乳动物细胞附着在蓝宝石盘上。我们的协议进一步涉及通过高压冻结,冻结固定和树脂嵌入的冷冻污染。然后,我们使用荧光光学显微镜在树脂包裹的样品中选择有丝分裂细胞。接下来是大规模电子断层扫描,以重建3D中所选的有丝分裂纺锤体。然后,生成和缝合的电子断层图用于半自动分段微管,以进行纺锤体组织的随后定量分析。因此,通过提供详细的相关光和电子显微镜(CLEM)方法,我们为细胞生物学家提供了一种工具集来简化纺锤体微管的3D可视化和分析(http://kiewisz.shinyapps.io/asga)。此外,我们指的是一个最近启动的平台,该平台允许交互式显示3D重建有丝分裂纺锤体(https://cfci.shinyapps.io/asga_3dviewer/)。

TD 2(DNA复制和特性传递)
- 在 ................................. 前期,核膜碎裂成碎片 - 在 ................................. 中期,纺锤体有丝分裂的赤道板形成 - 在 ................................. 中期,染色单体分离形成两组子染色体 - DNA 合成的时期称为 S 期 - 纺锤体有丝分裂由微管组成,微管是亚基微管蛋白的聚合物 - 染色体迁移是通过纺锤体微管与与每个染色体的着丝粒相关的结构结合实现的:着丝粒

迫使分裂的癌细胞死亡;低剂量药物组合防止纺锤体极聚集
摘要 有丝分裂在基于微管的纺锤体控制下,是抗癌治疗的一个有吸引力的靶点,因为癌细胞会经历频繁且不受控制的细胞分裂。破坏有丝分裂的微管靶向剂或有丝分裂激酶或微管马达的单分子抑制剂可以高效杀死癌细胞。然而,这些治疗方法存在严重的缺点:它们还针对经常分裂的健康组织,例如造血系统,并且由于原发性或获得性耐药机制,它们经常失去效力。在癌细胞分裂中出现的另一个目标是它们将有丝分裂纺锤体的极点“聚集”成双极结构的能力。这种机制对于癌细胞的特定存活是必要的,这些癌细胞由于经常存在异常的着丝粒数目或其他纺锤体缺陷而倾向于形成多极纺锤体。在这里,我们讨论了针对纺锤体极点聚集的组合治疗的最新发展,这些治疗专门针对具有异常着丝粒数目的癌细胞,并且由于其组合性质,有可能避免耐药机制。
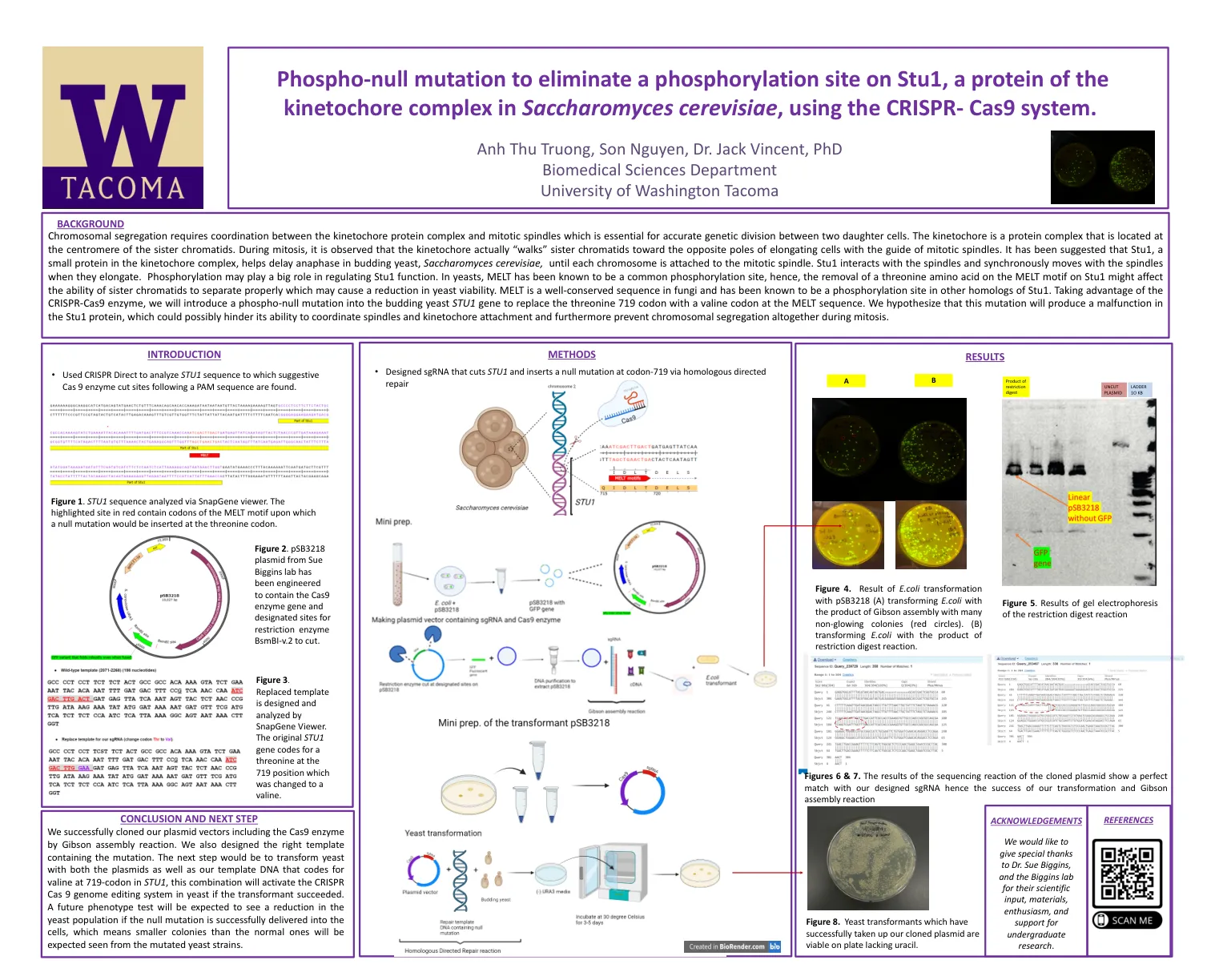
利用 CRISPR 技术进行磷酸化无效突变,消除酿酒酵母动粒复合体蛋白 Stu1 上的磷酸化位点
染色体分离需要动粒蛋白复合物和有丝分裂纺锤体的协调,这对于两个子细胞之间的准确遗传分裂至关重要。动粒是一种位于姊妹染色单体着丝粒的蛋白复合物。在有丝分裂过程中,可以观察到动粒实际上是在有丝分裂纺锤体的引导下将姊妹染色单体“引导”到伸长细胞的相反极点。有人提出,动粒复合物中的小蛋白 Stu1 有助于延迟芽殖酵母酿酒酵母的后期,直到每条染色体都附着在有丝分裂纺锤体上。Stu1 与纺锤体相互作用,并在纺锤体伸长时与其同步移动。磷酸化可能在调节 Stu1 功能方面发挥重要作用。在酵母中,MELT 是一种常见的磷酸化位点,因此,去除 Stu1 上 MELT 基序上的苏氨酸氨基酸可能会影响姐妹染色单体正确分离的能力,从而导致酵母活力下降。MELT 是真菌中保存良好的序列,并且已知是 Stu1 其他同源物中的磷酸化位点。利用 CRISPR-Cas9 酶,我们将在芽殖酵母 STU1 基因中引入磷酸化无效突变,以将 MELT 序列中的苏氨酸 719 密码子替换为缬氨酸密码子。我们假设这种突变会导致 Stu1 蛋白发生故障,这可能会阻碍其协调纺锤体和着丝粒附着的能力,并进一步阻止有丝分裂期间染色体分离。