XiaoMi-AI文件搜索系统
World File Search System线粒体

尿线粒体DNA 与原发性膜性肾病患者 ...
原发性膜性肾病 ( primary membranous nephro- pathy , PMN ) 是全球成人肾病综合征常见的病因 , 也是中国原发性肾小球疾病中发病率第二 、 增长 最快的疾病 [ 1 ] 。大多数 PMN 患者有典型的临床表 现 , 包括大量蛋白尿 、 低蛋白血症 、 水肿和高脂血 症等。近 30% 的 PMN 患者能够获得自发缓解 , 但 中危和高危患者 , 即大量蛋白尿 、 肾功能不稳定的 患者 , 缓解的可能性较低 [ 2 ] 。 既往研究表明 , 线粒体功能障碍在急性肾损伤 ( acute kidney injury , AKI ) 和慢性肾脏病 ( chronic kidney diseases , CKD ) 的发病机制和肾脏修复中发 挥关键作用 [ 3 - 4 ] 。线粒体功能与线粒体 DNA ( mito- chondrial DNA , mtDNA ) 的完整性密切相关 , 当线 粒体受损时 , mtDNA 会从线粒体基质释放到细胞 质或细胞外 , 进而激活氧化应激反应 , 并作为炎症 介质激活自然免疫炎症反应 [ 5 ] 。目前多项研究表 明 , 尿 mtDNA 是各种肾脏疾病中线粒体损伤的替 代标志物 [ 6 ] 。我们之前的研究表明 , mtDNA 在尿液 和肾脏组织中容易被检测到 , 其拷贝数与糖尿病肾 脏疾病的肾功能下降和肾脏病理结构改变有关 [ 7 ] 。 另一项研究指出 , 尿液中 mtDNA 与肾功能下降速 度有关 , 并能预测非糖尿病肾脏疾病患者血肌酐翻 倍或需要进行透析治疗的风险 [ 8 ] 。然而 , 尿 mtD- NA 在 PMN 患者中的改变及其对预后的预测作用 仍不明确。本研究旨在探讨尿 mtDNA 与 PMN 患
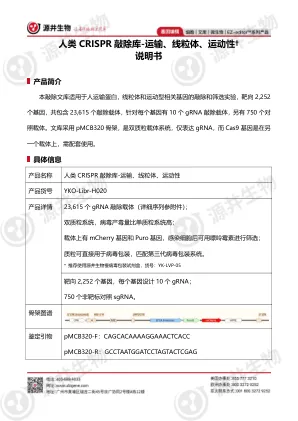

EZ-editor™ 人类CRISPR 敲除库-运输、线粒体、运动性†说明书
文库。除此之外,源井还提供CRISPR-KO、CRISPRa、CRISPRi 三大定制文库从高通量sgRNA 文

线粒体钙稳态作为线粒体医学的潜在目标
1. 引言. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ................. ... 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 。 83


线粒体离子通道
线粒体参与了多个细胞任务,例如ATP合成,代谢,代谢和离子转运,细胞凋亡的调节,线粒体DNA的发病,信号传导和遗传。线粒体的大多数正确功能基于大型电化学质子梯度,其成分(其内部线粒体膜电位)严格由通过线粒体内置的离子转运来控制。因此,线粒体功能严重取决于离子稳态,其干扰导致细胞功能异常。因此,发现通过膜影响离子通透性的线粒体离子通道定义了离子通道在不同细胞类型中的功能的新维度,这主要与线粒体离子通道在细胞生命和死亡中执行的重要任务有关。本综述总结了对动物线粒体离子通道的研究,特别关注其生物物理特性,分子身份和调节。此外,简要讨论了线粒体离子通道作为几种疾病的治疗靶标的潜力。


线粒体移植
线粒体移植是一种有前途的治疗方法,用于治疗由线粒体DNA突变引起的线粒体疾病,以及几种代谢性和神经性疾病。动物研究表明,线粒体移植可以改善细胞能量代谢,储存线粒体功能并防止细胞死亡。但是,需要解决挑战,例如将函数线粒体传递到体内正确的细胞,以及移植的线粒体的长期稳定性和功能。研究人员正在探索线粒体移植的新方法,包括使用纳米颗粒或CRISPR基因编辑。的基于移植线粒体的整合和功能的机制是复杂的,并且不完全理解,但是研究揭示了一些起作用的关键因素。在动物模型和人类试验中已经研究了线粒体移植的安全性和功效,但需要进行更多的研究来优化输送方法和评估长期的安全性和功效。使用线粒体移植的临床试验显示出不同的结果,突显了在该领域进行进一步研究的必要性。总而言之,尽管线粒体移植具有对各种疾病的治疗的巨大潜力,但仍需要更多的工作来克服挑战并评估其在人类试验中的安全性和功效。[BMB报告2023; 56(9):488-495]

