XiaoMi-AI文件搜索系统
World File Search System细胞表面

标题 经过基因编辑的 CD34+ 细胞源自人类 iPS 细胞(在体内但不是体外),可植入并分化为抗 HIV 细胞
iPS 细胞 | CCR5 | HIV 抗性 | 基因编辑 | 畸胎瘤 近期 HIV 研究的主要目标是开发一种“治愈”这种病毒感染的方法,避免终身接受抗逆转录病毒疗法 (ART)。实现这一目标的方法之一是删除或突变编码促进 HIV 感染和传播的蛋白质的基因。这一策略的一个有吸引力的候选基因是 Ccr5 基因,该基因突变导致 32 bp 缺失,已被证明与天然保护免受 HIV 感染和疾病有关 (1, 2)。Ccr5 基因编码 CCR5,这是一种人类细胞表面趋化因子受体,是 HIV 附着和感染细胞的辅助受体 (3, 4)。Ccr5 等位基因的 32 bp 缺失导致 CCR5 受体的截短异构体 CCR5 Δ 32,它不在细胞表面表达。因此,病毒进入细胞被阻止 (5)。诱导性多能干 (iPS) 细胞 (6) 能够分化为 CD34 + 造血干细胞 (HSC) (7),因此可以重建完整的免疫系统 (8, 9)。因此,这些 iPS 细胞是基因工程的首选目标。我们小组和其他小组已经证明,由健康个体 (10) 和接受 ART 治疗的 HIV 感染患者 (11) 的外周血单核细胞 (PBMC) 产生的 iPS 细胞可以经过基因编辑,使其 Ccr5 基因的野生型等位基因携带 Ccr5 Δ 32 突变 (12, 13)。值得注意的是,使用 CRISPR/Cas9 技术,可以修改 Ccr5 基因,使其具有与对 R5 嗜性病毒的抵抗力相关的天然 Δ 32 变体等位基因。此外,虽然截短的 CCR5 Δ 32 蛋白不存在于细胞表面,但它仍然表达,因此可能具有其他重要的生理作用(14-17)。我们已经证实,基因改造的 Ccr5 Δ 32 iPS 细胞可以在体外分化为 CD34 + HSC(10,18)。在适当的细胞培养条件下,它们可以产生各种
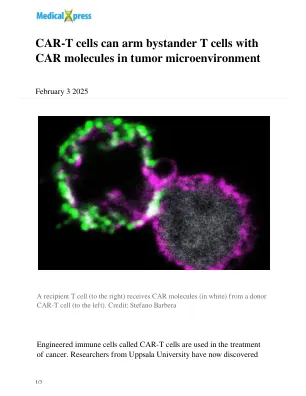
CAR-T 细胞可以在肿瘤微环境中用 CAR 分子武装旁观者 T 细胞
“我们的研究推翻了这一假设,我们证明,尽管需要细胞与细胞之间的接触,但特定分子不需要另一个细胞上的相应受体才能进行吞噬。相反,细胞表面分子周围的膜区域决定了它是否可以转移,”免疫学、遗传学和病理学系博士后研究员、该研究的第一作者兼通讯作者 Stefano Barbera 说。


Rhodotorula Mucilaginosa YR29能够累积PB
用六甲硅烷基处理的细胞已显示出某些细胞表面损伤,而不管真菌培养中使用的金属如何。尽管这可能是由于干燥过程引起的,这也会导致微胶囊的丢失(图5A-D)。 在此干燥过程中,处理的细胞在其拓扑结构没有变化。 仅在下部电子检测器(LED)进行PB处理时,揭示了非典型的三维泄漏(图。 5b)。 随后,在用Pb处理后,Cu和Zn可以在细胞表面观察到一些絮状物(图 5b-d)。 否则,从图。 E-P图的 5,观察到酵母菌保持其微胶囊,样品通过临界点过程(CPD)干燥。 微胶囊的方面是包围整个单元的薄层。 此外,此层5A-D)。在此干燥过程中,处理的细胞在其拓扑结构没有变化。仅在下部电子检测器(LED)进行PB处理时,揭示了非典型的三维泄漏(图。5b)。随后,在用Pb处理后,Cu和Zn可以在细胞表面观察到一些絮状物(图5b-d)。否则,从图。5,观察到酵母菌保持其微胶囊,样品通过临界点过程(CPD)干燥。微胶囊的方面是包围整个单元的薄层。此外,此层

肌动蛋白 - 膜连接器:合成重构系统的见解
在细胞表面,肌动蛋白细胞骨架和质膜在与细胞表面重塑有关的多种过程中相互相互作用。已知肌动蛋白细胞骨架可调节膜组织并重塑膜。为此,肌动蛋白 - 膜连接分子在调节肌动蛋白组装中起着重要作用,并在空间上指导肌动蛋白细胞骨架与膜之间的相互作用。虽然细胞中的研究为肌动蛋白 - 膜界面的分子组成和相互作用提供了丰富的知识,但复杂的分子相互作用使阐明界面处肌动蛋白 - 膜接头的精确效果变得具有挑战性。由模型膜和纯化蛋白组成的合成重构系统已成为一种强大的方法,可以阐明肌动蛋白 - 膜连接器如何直接肌动蛋白组装以驱动膜形状变化。在这篇综述中,我们将仅专注于使用重建系统研究的几个肌动膜连接器。我们将讨论这些重构系统的设计主要范围,以及它们如何为理解肌动蛋白 - 膜连接器的细胞功能做出贡献。最后,我们将在理解复杂的肌动蛋白 - 膜相互作用的未来研究方向上提供一个观点。
病毒蛋白 Nef 为癌症的现成 CAR-T 细胞疗法迈出了一步
显然,获胜者是一种名为 Nef 的蛋白质,HIV 病毒利用这种蛋白质逃避免疫系统的检测。它通过两种方式发挥作用:首先,它减少了 CAR-T 细胞表面一种名为 HLA-I 的蛋白质。HLA-I 通常充当免疫系统的红旗,发出信号表明存在问题并引发攻击——减少 HLA-I 有助于细胞不被发现。此外,Nef 有助于防止 CAR-T 细胞发生一种名为凋亡的细胞自杀形式。

评估培养基干细胞的质量特征
通过计算流式细胞仪的流动细胞来评估免疫标记,我们发现源自脂肪组织的中型组织干细胞显示出相对较好的表面烙印。具有烙印CD90,CD73,CD105的正比率分别为99.85%,99.34%和97.98%。这些表面标记的正比率往往高于Tanya Debnath的研究(CD90 98%,CD73 99%)。9的负标记,以2.06%的速度获得。 当负面制造商计算出CD34/45 0.2-2.5%,而HLADR为2.2%时,该指数往往与Tanya Debnath的研究相似。 9根据国际细胞治疗协会的2006年法规,中等干细胞必须显示某些细胞表面标志,例如CD73,CD90和CD105,并且不显示其他标志,包括表面分子CD45,CD34,CD14,CD14或CD11b,CD11b,CD79 Alpha或CD19和CD19和CD19和HLA-DR。 11这是胡椒9的负标记,以2.06%的速度获得。当负面制造商计算出CD34/45 0.2-2.5%,而HLADR为2.2%时,该指数往往与Tanya Debnath的研究相似。9根据国际细胞治疗协会的2006年法规,中等干细胞必须显示某些细胞表面标志,例如CD73,CD90和CD105,并且不显示其他标志,包括表面分子CD45,CD34,CD14,CD14或CD11b,CD11b,CD79 Alpha或CD19和CD19和CD19和HLA-DR。 11这是胡椒

信号通路下游至受体酪氨酸激酶
抽象的哺乳动物细胞具有调节细胞功能的各种不同细胞外刺激的能力。这通常涉及与细胞表面受体结合的配体以及随后的细胞内信号通路的激活。这些途径可能导致基因表达模式的变化,进而调节细胞生长,分化,迁移和功能。一种重要的细胞表面受体类型是受体酪氨酸激酶(RTK)。响应于配体结合的响应,rtks二聚,然后互相反磷酸化,从而导致下游途径的激活。虽然这些途径中的信号传导蛋白对于正常的细胞生长控制很重要,但如果不当调控它们可能导致不受控制的生长,有时甚至有时会导致癌症。因此,它们通常被认为是化学治疗药物药物靶标的良好候选者。RTK可以激活多个不同的信号通路。这些途径中的某些信号蛋白可以与其他RTK激活途径串扰,并且其中一些可以通过RTKS激活之外的多种机制激活。虽然RTK激活了各种不同的信号蛋白和途径,但在本综述中,我们将讨论包括MAPK途径,HER2/NEU途径,MTOR,MTOR和PAK激酶在内的几个关键途径的组件。我们概述了这些途径在细胞信号传导中的作用,并讨论如何将这些途径的不同组成部分视为癌症治疗的靶标。

化疗:药物 TZ 政策
Tafasitamab-cxix 是一种 Fc 修饰的单克隆抗体,可与表达于前 B 和成熟 B 淋巴细胞表面以及多种 B 细胞恶性肿瘤(包括弥漫性大 B 细胞淋巴瘤 (DLBCL))上的 CD19 抗原结合。与 CD19 结合后,tafasitamab-cxix 通过细胞凋亡和免疫效应机制介导 B 细胞裂解,包括抗体依赖性细胞毒性 (ADCC) 和抗体依赖性细胞吞噬作用 (ADCP)。在 DLBCL 肿瘤细胞中进行的体外研究中,与单独使用 tafasitamab-cxix 或来那度胺相比,tafasitamab-cxix 与来那度胺联合使用可提高 ADCC 活性。

全长层粘连蛋白蛋白对于干细胞至关重要
a)全长层粘连蛋白在组织上皮组织和内皮组织的基底膜方面起着核心作用。它们通过与几个细胞表面受体结合,包括整联蛋白,Syndecans,Lutheran血液组糖蛋白以及其他基质蛋白(如Nidogen和Agrin),从而激活细胞信号级联,从而形成细胞外基质和细胞之间的直接结合。b)层粘连蛋白521是天然干细胞生态位的钥匙基底膜蛋白,由人多能干细胞(HPSC)在预植物植入的胚胎的内部细胞质量中表达和分泌。[5] c)层粘连蛋白是三聚体蛋白,同工型是根据相互交织的α,β和γ链的组合指定的。