XiaoMi-AI文件搜索系统
World File Search System细胞表面

Q3 HCPCS I级和II级更新(2024年7月1日)
mepolizumab是白介素5(IL-5)拮抗剂(IgG1 kappa)。il-5是负责嗜酸性粒细胞生长和分化,募集,激活和存活的主要细胞因子。mepolizumab以100 pm的解离常数与IL-5结合,通过阻断在嗜酸性细胞表面表达的IL-5受体复合物的Alpha链的结合来抑制IL-5的生物活性。炎症是哮喘和嗜酸性肉芽肿的发病机理(EGPA)的重要组成部分。多种细胞类型(例如,肥大细胞,嗜酸性粒细胞,中性粒细胞,巨噬细胞,淋巴细胞)和介质(例如组胺,eicosanoids,白细胞素,细胞因子)参与炎症。mepolizumab,通过抑制IL-5信号传导,降低了嗜酸性粒细胞的产生和存活。但是,尚未确定哮喘和EGPA中的巨脂单抗作用的机制。

IHC面板标记
CD56,也称为神经细胞粘附分子,是一种在神经,神经胶质和骨骼肌细胞的细胞表面表达的均匀结合糖蛋白。CD56是天然杀伤细胞和许多其他免疫细胞的表型标记,包括α-β-beta T细胞,伽马三角细胞,树突状细胞和单核细胞。取决于蛋白质的剪接方式,功能可能会大大变化。通常,通过分析单个NCAM转录本发现了二十七种形式的NCAM。在发育中调节了NCAM的剪接形式的表达,因此提出了NCAM在发育中的作用。alpha 2,NCAM的8连锁多氨基酸修饰在神经发育和突触可塑性中起重要作用。在免疫组织化学中,CD56抗体可用于区分许多肿瘤,例如骨髓瘤,Wilm的肿瘤,神经母细胞瘤,Ewing的肉瘤等。

抗鼠CD44兔重组抗体,仅PBS
背景信息CD44是一种I型跨膜糖蛋白,该糖蛋白在胚胎干细胞上表达,并在其他细胞类型的各种水平上表达,包括结缔组织和骨髓。CD44表达在癌细胞的亚群中也被上调,并被公认为是癌症干细胞的分子标记(PMID:29747682)。它是一种细胞表面受体,通过其对透明质酸(HA)的亲和力以及可能通过其对其他配体的亲和力(PMID:10694938)介导细胞细胞和细胞矩阵相互作用。与HA的粘附在细胞迁移,肿瘤生长和进展中起重要作用。 CD44还参与淋巴细胞的激活,再循环和归巢,以及造血。与HA的粘附在细胞迁移,肿瘤生长和进展中起重要作用。CD44还参与淋巴细胞的激活,再循环和归巢,以及造血。

了解糖尿病的原因,症状和并发症
糖尿病被认为是危险因素,这主要是由于它在脂质代谢中引起的重大改变。糖尿病是由于缺乏胰岛素分泌或减少组织对胰岛素的组织感应性而引起的碳水化合物,脂肪和蛋白质的代谢受损的同型。该疾病的一个特征方面是胰岛素的有缺陷或不足的分泌反应,这表现在碳水化合物(葡萄糖)的不当利用中,因此高血糖。糖尿病之所以发生,是因为胰腺无法产生足够的激素胰岛素来满足人体的需求,或者由于这种激素无法正常工作(胰岛素抵抗)。 如果单个UAL在细胞中没有葡萄糖,则身体将从另一个来源(脂质)获得能量。 葡萄糖是胰腺从Langerhans胰岛的β细胞中释放胰岛素的主要信号。 细胞具有胰岛素受体,胰岛素与受体和mobi lizes葡萄糖转运蛋白(GLUT)结合,在脂肪组织中,它具有GLUT 4,在胰腺中,它具有GLUT 2。。 Gluts进入细胞表面并在细胞内传输葡萄糖。 大多数葡萄糖都进入糖裂解途径,其中大多数被转化为糖原(葡萄糖糖尿病之所以发生,是因为胰腺无法产生足够的激素胰岛素来满足人体的需求,或者由于这种激素无法正常工作(胰岛素抵抗)。如果单个UAL在细胞中没有葡萄糖,则身体将从另一个来源(脂质)获得能量。葡萄糖是胰腺从Langerhans胰岛的β细胞中释放胰岛素的主要信号。细胞具有胰岛素受体,胰岛素与受体和mobi lizes葡萄糖转运蛋白(GLUT)结合,在脂肪组织中,它具有GLUT 4,在胰腺中,它具有GLUT 2。Gluts进入细胞表面并在细胞内传输葡萄糖。大多数葡萄糖都进入糖裂解途径,其中大多数被转化为糖原(葡萄糖
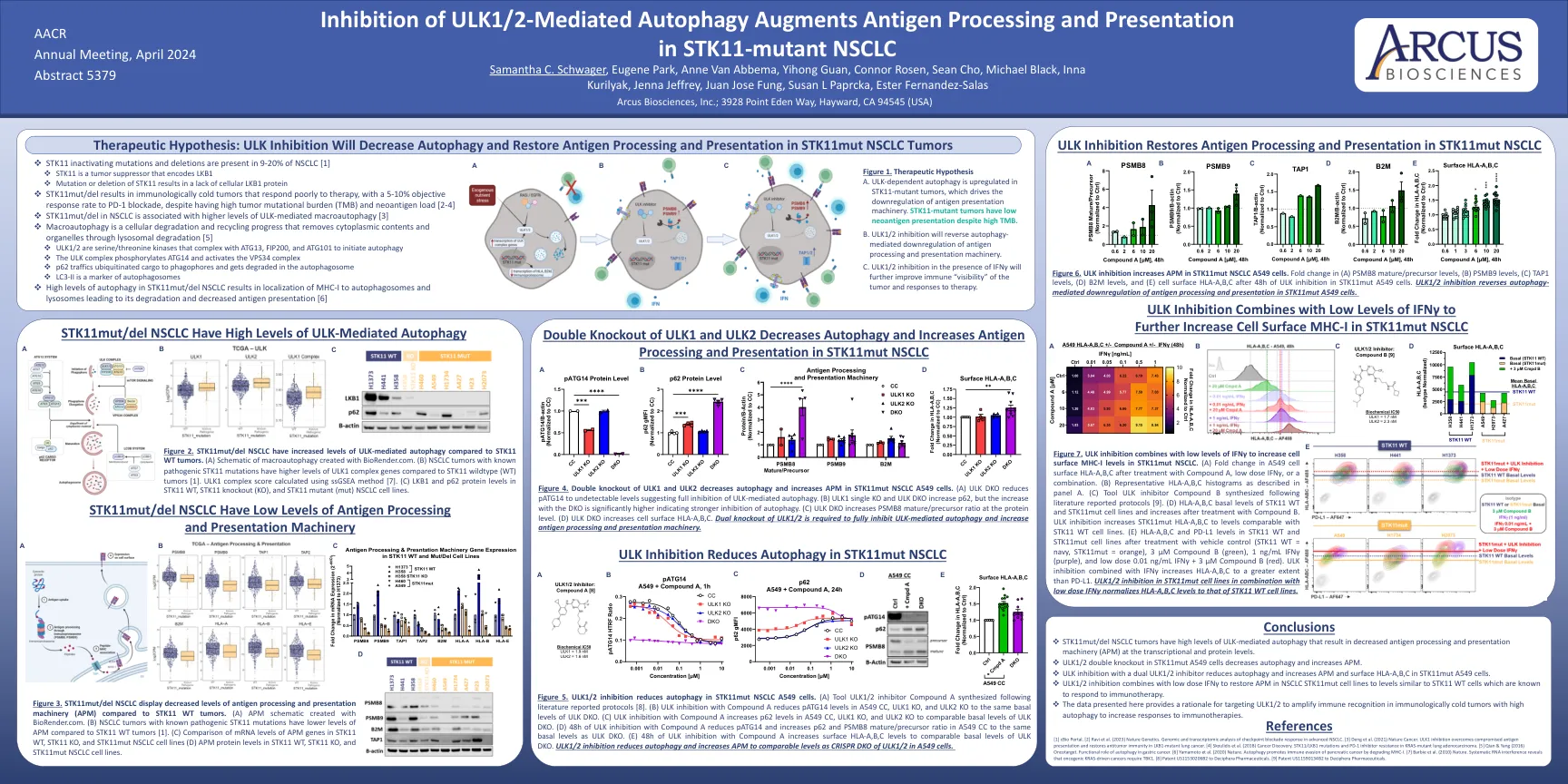
抑制ULK1/2介导的自噬增强抗原加工和表现 ARC-9:一项评估基于伊特鲁马德的治疗组合的随机研究
图2。与STK11 WT肿瘤相比, STK11MUT/DEL NSCLC具有ULK介导的自噬水平升高。 (a)用biorender.com创建的大噬细胞的示意图。 与STK11野生型(WT)肿瘤相比,具有已知致病性STK11突变的NSCLC肿瘤具有更高水平的ULK1复合基因[1]。 ULK1复合分数使用SSGSEA方法计算[7]。 (C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。 图4。 ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。 (a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。 (b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。 (c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。 (d)ULK DKO增加了细胞表面HLA-A,b,c。 需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。STK11MUT/DEL NSCLC具有ULK介导的自噬水平升高。(a)用biorender.com创建的大噬细胞的示意图。与STK11野生型(WT)肿瘤相比,具有已知致病性STK11突变的NSCLC肿瘤具有更高水平的ULK1复合基因[1]。ULK1复合分数使用SSGSEA方法计算[7]。 (C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。 图4。 ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。 (a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。 (b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。 (c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。 (d)ULK DKO增加了细胞表面HLA-A,b,c。 需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。ULK1复合分数使用SSGSEA方法计算[7]。(C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。图4。ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。(a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。(b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。(c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。(d)ULK DKO增加了细胞表面HLA-A,b,c。需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。

植物免疫受体途径作为对病原体的统一前沿
植物发展了先天免疫系统,以激活抗病性机制并抵御微生物入侵者。该系统包括由两类免疫受体引发的两个主要信号级联反应,即细胞表面免疫受体,也称为模式识别受体(PRRS)和细胞内免疫受体,也称为核苷酸结合结构域亮氨酸重复受体(NLR)。PRR和NLR具有不同的生化活性,并通过很大程度上独立的机制激活。但是,下游免疫反应和输出非常相似,表明两种途径之间的连通性和收敛性。的确,最近的研究显着提高了我们对两个cas虫之间相互依存与相互增强的亲密关系的理解。植物先天免疫的联合视图正在出现。

叶酸-叶酸受体作为药物载体在靶向给药系统中的研究进展
摘要 :靶向给药系统是肿瘤诊疗的有效方法,因其副作用小、疗效确切而受到广泛关注。叶酸受体在大多数癌细胞表面高表达,而在正常细胞表面低表达或不表达,且配体叶酸具有较高的亲和力,叶酸受体附着在药物载体上,可以靶向作用于癌细胞。本文介绍了叶酸和叶酸受体,简述了叶酸受体介导的靶向给药作用机制,讨论了叶酸偶联磁性纳米粒子、小分子叶酸的药物结合、叶酸受体结合蛋白、叶酸偶联多聚唾液酸四种叶酸-叶酸受体介导的肿瘤治疗进展,并分析了各治疗机制的优势点和未来发展趋势。

浸润性小叶乳腺癌情况说明书
激素(内分泌)疗法所有癌症都使用活检或手术后的乳腺组织进行检测,以确定它们是否对激素敏感(雌激素受体阳性或 ER+)。对激素敏感的癌症在细胞表面具有与雌性激素雌激素结合的受体,雌激素会刺激癌细胞生长。大多数侵袭性小叶癌都是雌激素受体阳性,这意味着您的医生可能会建议您进行激素疗法。激素疗法以不同的方式阻断激素对癌细胞的影响。有几种药物,您的医生会告诉您他们认为哪种药物最适合您。如果不存在雌激素受体(雌激素受体阴性或 ER-),可以进行测试以确定乳腺癌是否对另一种激素孕酮敏感。靶向疗法

了解 HLA 杂合性缺失以增强造血细胞移植后的治疗决策
流式细胞术在根据与正常造血骨髓群的差异识别异常群或微小残留病 (MRD) 方面发挥着至关重要的作用。异常群是通过细胞表面标志物 (CD 抗原) 的表达和抗原表达强度来识别的。Hematologics 的 ΔN:™(与正常的差异)流式细胞术侧重于表达强度而不是细胞频率来确定残留病。对于被视为异常的群,它必须是至少 40 个细胞或足够样本的 0.02% 的簇,并且异常与正常群至少有两个标准差。一旦根据 ΔN 确定细胞群为正常,就会利用免疫表型对母细胞进行分类。然后,这些分选细胞的 DNA 被发送到 Versiti 进行进一步的基因组分析。

用于体内人类基因组工程的可编程包膜运载载体
摘要:病毒和病毒衍生颗粒具有将分子递送至细胞的固有能力,但难以轻易改变细胞类型选择性,这阻碍了它们用于治疗递送。本文我们展示了通过展示在包裹 CRISPR-Cas9 蛋白和向导 RNA 的膜衍生颗粒上的抗体片段识别细胞表面标志,可以将基因组编辑工具靶向特定细胞。这些 Cas9 包装包膜递送载体 (Cas9-EDV) 用不同的展示抗体片段进行编程,在体外和体内混合细胞群中对靶细胞而不是旁观者细胞进行基因组编辑。该策略使得能够在人源化小鼠中生成基因组编辑的嵌合抗原受体 (CAR) T 细胞,从而建立了一种具有广泛治疗用途的新型可编程递送方式。