XiaoMi-AI文件搜索系统
World File Search System终末

特定癌症和糖尿病风险
21 世纪对公共健康的主要威胁之一是全球糖尿病患病率的上升 (1)。2 型糖尿病 (T2D) 是指导致 b 细胞衰竭的胰岛素抵抗和代偿性高胰岛素血症的初始阶段 (2)。2 型糖尿病的特征是慢性高血糖,随着时间的推移会损害终末器官 (2)。世界卫生组织报告称,每六例死亡中就有一例死于癌症,这使得癌症成为全球第二大死亡原因 (3)。癌症和糖尿病都是复杂的慢性疾病,给社会带来高昂的经济成本。这两种疾病在人体中同时发生已有 50 多年的报道 (4)。推测这两种疾病可能有相似的发展途径,例如免疫调节和细胞因子活性的失调 (5)。已发现肥胖、遗传易感性和接触某些环境因素等常见风险因素与癌症和糖尿病的发展有关 ( 5 , 6 )。鉴于已发现腹部脂肪会促进全身炎症,从而增加患癌症和糖尿病的风险,肥胖被认为是这两种疾病的潜在原因之一 ( 6 )。流行病学证据表明,2 型糖尿病患者更容易患上多种恶性肿瘤 ( 7 )。例如,糖尿病显著增加肝癌和胰腺癌 (PC) 的相对风险 ( 8 , 9 ),但对于其他癌症的证据较少。由于某些恶性肿瘤的发展可能先于 2 型糖尿病并导致 2 型糖尿病,因此还应考虑这两种疾病的潜在反向因果关系。例如,胰腺癌可能会促进 2 型糖尿病的发展 ( 10 )。韩国最近的一项研究表明,癌症会增加癌症幸存者患糖尿病的风险,与传统的糖尿病风险因素无关 ( 11 )。糖尿病风险在癌症确诊后的前两年最为显著,并且风险升高的趋势可持续长达 10 年 ( 11 )。此外,循环细胞因子通过促进胰岛素抵抗和增加肝糖异生而加剧癌症患者的高血糖症 ( 12 )。PC 的标准肿瘤标志物是较高水平的 CA19-9,血清 CA19-9 水平升高与血糖调节不足的严重程度有关 ( 13 , 14 )。有研究表明,癌症治疗的幸存者在其余生中患糖尿病和代谢综合征等内分泌疾病的风险更高 ( 15 )。例如,最近的研究表明,糖尿病更有可能

靶向 CD96 可通过增强宫颈癌中的 CD8+ TIL 功能来克服 PD-1 阻断耐药性
摘要 背景 复发性和晚期宫颈癌 (CC) 的预后仍然很差,因此需要新型疗法来治疗。尽管针对程序性细胞死亡蛋白 1 (PD-1) 通路的疗法已被批准用于治疗 CC,但很大一部分患者表现出先天性耐药性。联合使用检查点抑制剂可以增强其疗效。方法 从 CC 患者获取血液样本、肿瘤标本和瘤周 (PT) 组织。通过流式细胞术分析 CC 标本中 CD8+ T 细胞的抑制性受体表达和表型分析。通过免疫组织化学和免疫荧光测量肿瘤细胞表达的 CD96 配体。通过基于 CC 标本单细胞培养的离体治疗试验评估对派姆单抗的敏感性。使用离体治疗试验和人乳头瘤病毒阳性 TC-1 异种移植小鼠模型在体内探索 PD-1 和/或 CD96 阻断的疗效。结果我们发现对 PD-1 阻断不敏感的 CC 患者的 CD8+ 肿瘤浸润淋巴细胞 (TIL) 上 CD96 表达升高。这些表达 CD96 的 CD8+ TIL 通常共表达 PD-1。来自 scRNA-seq 数据的 CD96+CD8+/CD96−CD8+ T 细胞基因特征的比例与宫颈鳞状细胞癌和宫颈腺癌患者的不良生存率显着相关。与血液和 PT 组织相比,与 CD96 竞争的共刺激受体 CD226 在肿瘤中下调。CD96 和具有 Ig 和 ITIM 结构域的 T 细胞免疫受体 (TIGIT) 在肿瘤内 CD8+ T 细胞上上调。CD226/CD96/TIGIT 信号配体在 CC 肿瘤组织中广泛表达。表型分析显示,PD-1+CD96+CD8+ TIL 表现出终末衰竭效应表型,T 细胞免疫球蛋白黏蛋白受体 3 (TIM-3) 和颗粒酶 B (GZMB) 水平高,促炎细胞因子和细胞毒性分子水平极低。PD-1+CD96 细胞表现出前体衰竭表型,TCF-1 阳性。PD-1 阻断后,CD8+ TIL 进一步上调 CD96。CD96 阻断治疗显著增强了 PD-1 阻断,从而抑制肿瘤生长,并改善小鼠和 CC 标本模型中 CD8+ TIL 的功能。

双特异性免疫细胞参与者在体外增强了CD16+ NK细胞和巨噬细胞的抗癌活性,并消除了NK人源化NOG小鼠的癌症转移
抽象背景在肿瘤微环境中发现了多种类型的产生IL-17的免疫细胞。然而,它们在CD8 +肿瘤浸润淋巴细胞(TILS)的肿瘤进展和精疲力尽中的作用尚不清楚。确定17型免疫力在肿瘤中的作用的方法,我们研究了B16F10黑色素瘤的生长以及CD8 + TIL在IL17A - / - 小鼠中的耗尽,IL17A CRE R26 DTA小鼠,RORγT抑制剂造成的小鼠,或其相应的对照组。在B16F10的先天小鼠中进行了肿瘤特异性IL-17产生的T细胞的过继转移。抗CD4或抗Ly6g抗体分别用来耗尽CD4 + T细胞或CD11b + Gr-1 HI髓样细胞的体内。通过询问TCGA数据集,评估了人类癌症中17型免疫力与T细胞衰竭之间的相关性。结果CD4 + T细胞的耗竭促进了CD8 + T细胞的耗尽,而肿瘤中产生IL-17的CD8 + T(TC17)细胞的耗尽。与产生CD8 + T(TC1)细胞不同,肿瘤浸润的TC17细胞表现出CD103 + KLRG1-IL-7RαHIHI组织驻留的记忆样表型,并且细胞溶解度不佳。产生IL-17的肿瘤特异性T细胞的产物转移增加,而产生IL-17的细胞的耗竭降低,肿瘤中PD-1 HI TIM3 + TOX +终末耗尽的CD8 + T细胞的频率。封锁IL-17或RORγT途径抑制CD8 + T细胞的耗尽,还会延迟体内肿瘤的生长。封锁IL-17或RORγT途径抑制CD8 + T细胞的耗尽,还会延迟体内肿瘤的生长。与这些结果一致,人类TCGA分析揭示了多种癌症中17型和CD8 + T细胞耗尽签名基因集之间存在很强的正相关。结论IL-17产生的细胞促进了CD8 + T细胞的末端衰竭和体内肿瘤进展,可以通过IL-17或RORγT途径的阻断来逆转。这些发现揭示了IL-17-产生细胞作为促进CD8 + T细胞耗尽的肿瘤细胞的新作用,并提出17型免疫力作为癌症免疫疗法的有希望的靶标。

SGLT-2 抑制剂作为二甲双胍的附加治疗对 2 型糖尿病患者 P 波指数和心房电机械的影响
缩写:LA,左心房;LAVI,左心房容积指数;IEMD,心房间电机械延迟;LEMD,左侧心房内电机械延迟;LVEF,左心室射血分数;PA,P 至 A' 间期;PTFV1,V1 中的 P 终末力;PWD,P 波弥散;REMD,右侧心房内电机械延迟。 *I 组 vs. III 组;p = .035。**I 组 vs. III 组 p = .002。***I 组 vs. II 组;p = .042,I 组 vs. III 组;p < .001,II 组 vs. III 组;p = .033。****I 组 vs. II 组;p = .011,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .005。*****I 组 vs. II 组; p = .035,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .005。+ I 组 vs. II 组;p = .042,I 组 vs. III 组;p < .001,II 组 vs. III 组;p = .021。++ I 组 vs. II 组;p = .012,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .004。+++ I 组 vs. II 组;p = .002,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p < .001。¥ I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .038。¥¥ I 组 vs. II 组;p = .026,I 组 vs. III 组; p < .0001,II 组与 III 组;p = .023。¥¥¥ I 组与 II 组;p = .016,I 组与 III 组;p < .0001,II 组与 III 组;p = .036。§ I 组与 II 组;p = .042,I 组与 III 组;p < .001,II 组与 III 组;p = .039。§§ I 组与 II 组;p = .044,I 组与 III 组;p = .003,II 组与 III 组;p = .040。

RNA聚合酶II暂停暂时协调细胞周期进程和红细胞分化 Danya J. Martell 1,2,3,4 , Hope E. Merens 1 , C
1 哈佛大学遗传学系,马萨诸塞州波士顿,2 哈佛医学院波士顿儿童医院血液学/肿瘤学分部,马萨诸塞州波士顿,3 哈佛医学院丹娜法伯癌症研究所儿科肿瘤学系,马萨诸塞州波士顿,4 麻省理工学院和哈佛大学布罗德研究所,马萨诸塞州剑桥,5 米兰大学临床科学与社区系,IRCCS Ca'Granda Foundation Maggiore Policlinico 医院,意大利米兰,6 牛津大学 MRC Weatherall 分子医学研究所,英国牛津,7 牛津大学 MRC Weatherall 分子医学研究所 MRC 分子血液学部,英国牛津,8 牛津大学医院 NHS 基金会信托牛津遗传学实验室,英国牛津,9 NIHR 牛津生物医学研究中心和 BRC/NHS 转化分子诊断中心,约翰拉德克利夫医院,英国牛津,10 牛津大学医院 NHS 基金会血液学系Trust,牛津,英国 # 通讯作者 摘要 启动子近端暂停的 RNA 聚合酶 II (Pol II) 的控制释放进入生产性延长是基因调控的重要步骤。然而,对 Pol II 暂停进行功能分析很困难,因为调节暂停释放的因素几乎都是必需的。在这项研究中,我们在与 HBB 突变无关的 β-地中海贫血患者中发现了 SUPT5H(编码 SPT5)的杂合功能丧失突变。在健康人类细胞的红细胞生成过程中,细胞周期基因在从祖细胞到前体的转变中高度暂停。当通过 SUPT5H 编辑重现致病突变时,Pol II 暂停释放被全面打乱,从祖细胞到前体的转变被延迟,其特点是红细胞特异性基因表达和细胞周期动力学出现短暂滞后。尽管存在这种延迟,细胞仍会终末分化,细胞周期相分布也会恢复正常。因此,阻碍暂停释放会扰乱红细胞生成过程中关键转变处的增殖和分化动力学,揭示了 Pol II 暂停在细胞周期和分化之间的时间协调中的作用。 简介 细胞分化是一个严格调控的多步骤过程,需要细胞类型特异性基因表达程序中的几种转变。因此,转录的精确调控是控制基因表达的关键步骤,是细胞分化的基础 (Young 2011; Lee and Young 2013; Jonkers and Lis 2015)。RNA 聚合酶 II (Pol II) 暂停

钙结合蛋白 S100P 促进肿瘤进展但增强乳腺癌化学敏感性
乳腺癌是女性最常见的恶性肿瘤,也是癌症死亡的第二大原因 (1)。化疗被广泛用于提高乳腺癌患者的生存率。然而,一些患者不可避免地会以内在或获得性的方式表现出化学耐药性。为了更好地治疗这些乳腺癌患者,解决这个问题是一个巨大的挑战。对于三阴性乳腺癌 (TNBC) 类型或转移性患者尤其如此,因为对化疗药物的反应可能是他们生存的关键 (2)。因此,在开发更好的治疗方法来满足临床需求之前,揭示化学耐药背后的复杂机制是先决条件。S100 家族由 20 多个小的二聚体 Ca 2 + 结合蛋白组成,是螺旋-环-螺旋 (EF-hand) 超家族中最大的类群 (3)。研究发现,S100 蛋白参与调节钙稳态、细胞增殖和凋亡、细胞侵袭和运动、癌症转移、血管生成、细胞骨架相互作用、蛋白质磷酸化、转录因子调节、自身免疫、趋化性和炎症 ( 4 )。S100P 最初是在人胎盘中发现的 ( 5 ),也在胃、膀胱和骨髓等其他器官中表达 ( 6 )。它含有一个称为 EF-手形基序的特征结构域,以细胞内或分泌的同型或异型二聚体形式存在,其组成取决于细胞环境 ( 7 )。 S100P 的功能主要归因于它与调节肌动蛋白细胞骨架动力学和细胞外基质重塑的几种分子相互作用或对其进行调节,包括 Ezrin、IQGAP1、肌球蛋白 IIA、组织蛋白酶 D 和磷酸化肌动蛋白 (8-11)。S100P 在细胞内成分和细胞外基质中充当信号分子 (12,13)。虽然调节 S100P 的机制尚未完全证实,但 DNA 微阵列表明雌二醇 (14)、孕酮 (15) 和 HER2 过表达 (16) 会上调 S100P,这与临床研究结果一致,即高 S100P 水平与 ER/PR 和 HER2 过表达肿瘤有关。 S100P 与 ER 表达之间的显著关联意味着 S100P 参与了乳腺癌发生的早期阶段(17)。S100P 还与乳腺上皮细胞体外永生化、肿瘤进展和患者早期复发有关(17-19)。S100P 最近因其在恶性转化和肿瘤进展中的作用以及在预测几种癌症类型的预后和转移中的作用而引起了广泛关注(20)。S100P 在乳腺癌发生和进展中的作用也有报道(17,19)。S100P 在 TNBC 组织中表达升高 (21),与 TNBC 患者的生存率低有关 (22)。研究表明,与其他 TNBC 患者相比,S100P 和 Ezrin 细胞质水平较低的 TNBC 患者的无病生存率 (DFS) 更高 (23)。S100P 被认为通过激活晚期糖基化终末受体发挥其致癌活性

登革热和奥罗普切病毒感染风险增加
费城公共卫生局 (PDPH) 正在通知该地区的医疗保健提供者,由于登革热或奥罗普切病毒、具有相似临床表现和重叠爆发区域的虫媒病毒感染,旅行相关感染的风险增加。2024 年,美洲和波多黎各的国家继续创纪录地爆发登革热病毒。与此同时,奥罗普切病毒感染有所增加,从亚马逊盆地的流行地区蔓延到新的地区,包括巴西、玻利维亚、秘鲁、哥伦比亚、多米尼加共和国和古巴。流行病学和临床特征:登革热:登革热是全球最常见的虫媒病毒疾病。登革热病毒通过受感染的伊蚊叮咬传播,其中埃及伊蚊是最常见的媒介。该物种通常生活在热带和亚热带气候中,并在美国大陆南部和西南部地区发现,这些地区发生了有限的局部疫情。目前,佛罗里达州和加利福尼亚州洛杉矶已报告登革热的局部传播。在费城,白纹伊蚊是传播登革热的效率低得多的媒介,在蚊子肆虐的季节里随处可见。登革热是由四种不同但密切相关的登革热病毒 (DENV-1、-2、-3、-4) 引起的,因此有登革热病毒病史的人有可能感染不同类型的病毒。大约 75% 的登革热感染是无症状的,但剩余四分之一的人有症状,症状可从轻微到严重不等。患者通常表现为发烧,并伴有恶心、呕吐、皮疹、肌肉疼痛、严重骨痛、头痛或白细胞计数低。大约 5% 的有症状登革热患者会发展为严重疾病,其特征是严重出血、休克、呼吸窘迫或终末器官衰竭。1 岁以下的婴儿、孕妇、老年人和患有某些疾病的人感染严重登革热的风险更高。奥罗普切病毒:奥罗普切病毒是南美洲和加勒比地区一种新出现的节肢动物传播病毒,由蠓和某些蚊子传播。虽然奥罗普切病毒已经传播到亚马逊盆地以外的新地区,但截至 2024 年,美国仅报告了来自 5 个州的 74 例旅行相关感染。奥罗普切病毒的症状类似于其他虫媒病毒疾病,包括登革热。大多数有症状的患者有自限性发热性疾病,伴有肌痛、头痛和关节痛。其他症状包括眶后疼痛、畏光、呕吐、腹泻、疲劳、斑丘疹、结膜充血和腹痛。临床实验室检查结果可能包括淋巴细胞减少和白细胞减少、C 反应蛋白 (CRP) 升高和肝酶略有升高。孕妇、老年人和患有潜在疾病的人(例如免疫抑制、高血压、糖尿病、患有心脏病或心血管疾病的人患严重疾病的风险更大,并发症包括出血表现(例如鼻出血、牙龈出血、黑便、月经过多、瘀点)和神经侵袭性疾病(例如脑膜炎、脑膜脑炎)。已报告两例死亡病例。可能的垂直传播与
社论
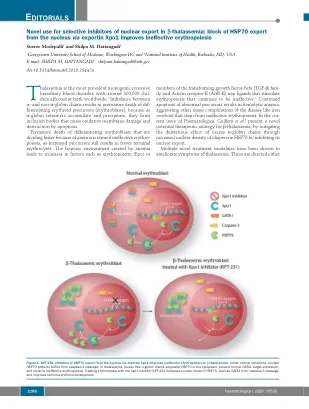
改善未结合的A -Globin和非A -Globin链之间的平衡或纠正无效的红细胞。修饰的TFG-β家族受体拮抗剂,如Sotatercemp(ACE-011)和Luspatercept(ACE-536)阻止配体与ACTR-II受体结合,并随后激活SMAD4信号通路,4改善Ery-Throid Throid Cell和红细胞的产生。通过CRISPR Therapeatics成功的基因疗法实现了未结合A -Globin与非A -Globin链的异常比例,并得到了波士顿顶点药物的支持。称为CTX001的体细胞疗法使用了编辑的患者自己的造血干细胞(HSC)来刺激胎儿血红蛋白的产生。5通过XPO1抑制HSP70的细胞内局部局部局部局部可能会合并这两个治疗目标。几条证据表明,红细胞使用分子伴侣在红细胞发育过程中对不稳定的过量A -Glo- bin链分割,6-8,因此,靶向这种伴侣的靶向时,当过量的globobin tetramer会累积时,靶向这种伴侣可能在β -tha -thaplamasemia中有用。许多组指出,分子伴侣HSP70在红细胞9-11中积聚至高水平,对于简化胚芽成熟很重要。11正常的人红细胞成熟需要在成熟后期的caspase-3瞬时激活,以防止过度的红细胞生产。激活的胱天蛋白酶可以切割GATA-1,从而导致成熟停滞和/或凋亡。12 Ribeil等。 14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。12 Ribeil等。14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。表明EPO会导致HSP70转移到核中,结合GATA-1并保护其免受caspase-3裂解。相反,在EPO剥夺期间,HSP70被排除在细胞核中,而GATA-1被cas-pase-3裂解,导致凋亡死亡。13因此,HSP70的细胞内位置的改变似乎在红细胞生存力中起关键作用(图1)。在β-丘脑贫血中观察到的无效性红细胞生成的特征是在多染色体阶段加速了红细胞分化,成熟停滞和凋亡。在人β-thal虫蛋白粒细胞的成熟过程中,HSP70直接通过过量的游离A-格珠蛋白链直接在细胞质中螯合(图1)。核定靶向的HSP70突变体或caspase-3-无分解的GATA-1突变体恢复了β-thal核阿无血成红细胞的终末成熟。14在Haematologica,Guillem等。3跟进这种机制,以表明导出蛋白1(XPO1)调节在正常条件下HSP70在红细胞中的HSP70的核质质位置。Guillem等。证实,用XPO1抑制剂KPT-251治疗红细胞增加了HSP70的核水平,从caspase-3裂解中救出了GATA1,并改善了末端红细胞原理(图1)。尽管使用核出口的选择性抑制剂(SINE)用于治疗淋巴瘤和多骨髓瘤

人胎盘中合成肌细胞细胞谱系发育的关键调节剂
背景。胎盘是一种瞬态器官,在怀孕期间形成以支持胎儿发育并调节影响慢性疾病风险的环境线索的暴露。胎盘在许多方面支持胎儿发育,包括促进营养和氧气交换,去除有害废物产品,产生关键的激素(例如人类绒毛膜促性腺激素)以及提供免疫保护。这些功能在很大程度上是由被称为合胞素细胞和额外滋养细胞细胞的终末分化的滋养细胞执行的。尽管合成肌细胞细胞和跨性滋养细胞细胞的重要性,但仍不清楚它们如何专门支持最佳胎儿发育。目标。使用功能方法丧失来确定合成肌细胞细胞谱系发育的转录调节因子。方法。候选转录因子(TBX3,VGLL3和ATF3)使用慢病毒介导的短发蛋白RNA(SHTBX3,SHTBX3,SHVGLL3或SHATF3)使用胞质衍生的人滋养细胞干细胞中击倒。将非特异性shRNA(SHCONTROL)用作对照。转导后,使用紫霉素选择细胞,并分别通过RT-QPCR和Western印迹在转录本和蛋白质水平上确认敲低效率。通过功能和转录组评估评估了转录因子敲低对滋养细胞干细胞分化为合成型肉芽细胞的影响。结果。结论。未来的方向。与用SHControl转导的细胞相比,SHTBX3和SHVGLL3的转导在合成型细胞细胞分化后导致形态异常。 可以使用滋养细胞干细胞中的功能方法丧失来评估候选转录调节剂对合成细胞细胞谱系发育的关键贡献。 初步结果表明,TBX3和VGLL3对于建立合成型细胞细胞谱系至关重要。 然而,需要更深入的表征来识别TBX3和VGLL3调节合成细胞成分的发育的分子机制。 未来的研究将包括完成剩余的候选转录因子,ATF3,全基因组评估(例如ATAC-SEQ)的shRNA敲低,以及所有SHRNA转换的其他功能输出,例如人类绒毛膜促性腺激素的产生。在合成型细胞细胞分化后导致形态异常。可以使用滋养细胞干细胞中的功能方法丧失来评估候选转录调节剂对合成细胞细胞谱系发育的关键贡献。初步结果表明,TBX3和VGLL3对于建立合成型细胞细胞谱系至关重要。然而,需要更深入的表征来识别TBX3和VGLL3调节合成细胞成分的发育的分子机制。未来的研究将包括完成剩余的候选转录因子,ATF3,全基因组评估(例如ATAC-SEQ)的shRNA敲低,以及所有SHRNA转换的其他功能输出,例如人类绒毛膜促性腺激素的产生。

病毒疾病中的反应性星形胶质细胞:朋友还是敌人?
在健康的大脑中,星形胶质细胞在神经元传播和血液 - 脑屏障(BBB)完整性中起着至关重要的作用。星形胶质细胞向反应状态的转化构成了中枢神经系统(CNS)对侮辱和大脑环境变化的生物学反应。众所周知,星形胶质细胞可以独立于神经元复制和积累王子[1-5]。然而,对它们的反应性转移对神经元功能和神经变性的影响知之甚少。在prion疾病中,反应性星形胶质细胞的有益作用似乎与星形胶质细胞生产的牛奶脂肪球表皮生长因子8(MFGE8)有关,这促进了凋亡人体的吞噬和细胞脱布的清除[6]。然而,在评估反应性星形胶质细胞对疾病进展的总体影响时,在保护稳态角色的潜在缺陷和有害功能的出现之外,至关重要的是,至关重要。最近的研究表明,反应性星形胶质细胞可能对神经元和内皮细胞产生净负面影响。从受prion感染的动物中分离出的反应性星形胶质细胞对原发性神经元表现出不利影响,导致树突状脊柱大小和密度降低以及突触完整性的损害[7](图1)。突触毒性作用是通过星形细胞分泌组的变化介导的,突出了信号传导途径在神经元功能障碍中的潜在作用。除了对神经元的影响外,反应性星形胶质细胞破坏了BBB的完整性。共培养实验涉及来自病毒感染的动物的星形胶质细胞或暴露于反应性星形胶质细胞的条件培养基中,诱导了从正常小鼠分离的内皮细胞中与疾病相关的表型[8](图1)(图1)。这种表型通过紧密和粘附连接蛋白的下调和异常定位以及内皮层的渗透性提高来征收这种表型。值得注意的是,星形胶质细胞激活程度和与prion疾病的孵育时间之间观察到非常强的反向相关[9]。具有快速疾病进展的动物群体表现出更严重的天线反应性,这表明星形胶质细胞的表型变化与缓解严重程度之间存在潜在的联系。这种观察结果提出了反应性星形胶质细胞的表型变化有助于更快的疾病进展的可能性。与这一假设一致,通过选择性靶向PERK信号传导的反应性星形胶质细胞中未折叠的蛋白反应的抑制作用,可以将其延长到小鼠中终末疾病的孵育时间[10]。总而言之,与Prion疾病相关的反应性星形胶质细胞对神经元和内皮细胞表现出有害的影响,并且可能是导致疾病进展的因素。阐明驱动星形胶质反应性的基本机制可能具有减轻与Prion疾病相关的神经退行性过程的治疗潜力。