XiaoMi-AI文件搜索系统
World File Search System结构域

与构象相关的自由能概况...
KRAS基因G12突变与多种癌症有关。采用多重复制高斯加速分子动力学(MR-GaMD)模拟研究了G12C、G12D和G12R突变引起的开关结构域构象变化。自由能图表明,与GTP结合的WT KRAS相比,G12C、G12D和G12R诱导的能量状态更高,使开关结构域的构象更加无序,从而干扰KRAS与效应分子的结合。基于MR-GaMD轨迹的动力学分析表明,G12C、G12D和G12R不仅改变了开关结构域的灵活性,而且影响了其运动行为,这表明这三个突变可用于调控KRAS的活性。相互作用网络分析验证了GTP与开关S Ⅰ相互作用的不稳定性在开关结构域的高度无序状态中起着重要作用。此项工作有望为深入了解KRAS的功能提供有用的信息。

NLRP3炎症小体激活、代谢危险信号......
NOD 样受体家族含吡啶结构域 3 (NLRP3) 炎症小体是一种寡聚复合物,可响应病原体感染的外源信号和非微生物来源的内源性危险信号而组装。当 NLRP3 炎症小体组装激活 caspase-1 时,它会促进炎症细胞因子白细胞介素-1B 和 IL-18 的成熟和释放。NLRP3 炎症小体的异常激活与各种疾病有关,包括慢性炎症、代谢和心血管疾病。NLRP3 炎症小体可以通过几种主要机制激活,包括 K + 外排、溶酶体损伤和线粒体活性氧的产生。有趣的是,代谢危险信号会激活 NLRP3 炎症小体以诱发代谢疾病。 NLRP3 包含三个关键结构域:N 端吡啶结构域、中央核苷酸结合结构域和 C 端富含亮氨酸重复结构域。蛋白质-蛋白质相互作用充当“踏板或刹车”,控制 NLRP3 炎症小体的激活。在这篇综述中,我们介绍了代谢危险信号诱导后或通过与 NLRP3 的蛋白质-蛋白质相互作用(可能发生在代谢疾病中)激活 NLRP3 炎症小体的潜在机制。了解这些机制将有助于开发治疗 NLRP3 相关代谢疾病的特定抑制剂。

Epimark甲基化的DNA富集套件,E2600,手册
通过与人MBD2A蛋白的甲基-CPG结合结构域结合与人IgG1(MBD2A-FC的FC尾巴)的甲基-CPG结合结构域结合,将甲基化的DNA从碎片的基因组DNA(5 ng-25μg)中分离出来,该蛋白与人IgG1(MBD2A-FC)的FC尾部结合,从而与paramagnetic Hydophilic Protein a betein a bead a bead a bead a bead a bead a bead a bead a bead a bead a bead a。两个FC结构域可以与具有高亲和力的蛋白质A上的一个位点结合(K d = 10 –7)。由于FC片段是二聚体,因此四个MBD2结构域暴露于每个分子蛋白A分子的溶剂,从而增加了相对平衡常数100倍。这种稳定的复合物将选择性地结合含有DNA的双链甲基化的CpG。在简单的洗涤步骤随后进行磁捕获后,通过在65°C下孵育,富集的DNA样品很容易在少量无核酸酶的水中洗脱。样本可以立即通过多种方法进行下游分析,包括:

SP120的选择性DNA结合(人HNRNP U的大鼠直系同源物)是由精氨酸 - 甘氨酸富含结构域介导的,并由RNA
人类蛋白质异质核糖核蛋白U(HNRNP U)也称为支架附着因子A(SAF-A)及其直系同源大鼠蛋白SP120是丰富的多功能核蛋白,可直接与DNA和RNA结合。富含精氨酸和甘氨酸的HNRNP U的C末端区域对于与RNA的相互作用至关重要,而SAF-A称为SAP结构域的N末端区域已归因于DNA结合。我们报告说,大鼠HNRNP U特异性和合作结合了称为核支架/基质相关区域(S/MAR)的富含的DNA,尽管其详细机制尚不清楚。在本研究分析中,HNRNP U缺失突变体首次揭示了富含arg-gly的C末端结构域(此处定义为“ RG结构域”)对于S/MAR-MAR-MAR-MAR-SELECHECTive DNA结合活性至关重要。rg域单独与S/MAR直接结合,并与SAP结构域共存具有协同作用。结合被Netropsin抑制,Netropsin是一种次要的凹槽粘合剂,偏爱富含S/MAR的成对,这表明RG结构域与S/MAR DNA的小凹槽相互作用。有趣的是,过量的RNA减弱了HNRNP U.综上所述,HNRNP U可能是RNA调节的S/MAR DNA识别的关键元素,从而有助于染色质区室的动态结构变化。

自身炎症疾病的异同......
缩写:ALP,碱性磷酸酶;ASC,含 CARD 的凋亡相关斑点样蛋白;CARD,胱天蛋白酶活化和募集结构域;CAPS,冷热蛋白相关周期性综合征;CINCA,慢性婴儿神经皮肤关节综合征;DAMPs,危险相关分子模式;DLBCL,弥漫性大 B 细胞淋巴瘤;ESR,红细胞沉降率;FCAS,家族性冷自发炎综合征;GSDMD,胃蛋白酶 D;IL-1R。IL-1 受体,IL-1RA;IL-1 受体拮抗剂,MGUS;意义不明确的单克隆丙种球蛋白病,MWS;马克-韦尔斯综合征,MYD88;髓系分化原发反应基因 88,NLR;NOD 样受体,NLRC4; NLR 家族胱天蛋白酶募集结构域含 4,NLRP3;NLR 家族,含吡啶结构域 3;NOD,核苷酸结合寡聚化结构域;NOMID,新生儿发病多系统炎症疾病;PGA,医生整体评估;

在 CRISPR-Cas12a 中,α-螺旋盖引导目标 DNA 进行催化
CRISPR-Cas12a 是一种强大的 RNA 引导基因组编辑系统,它利用其单个 RuvC 核酸酶结构域通过顺序机制产生双链 DNA 断裂,其中非靶链的初始切割随后是靶链切割。目前尚不清楚空间上相距甚远的 DNA 靶链如何向 RuvC 催化核心移动。在这里,连续数十微秒的分子动力学和自由能模拟表明,位于 RuvC 结构域内的 α 螺旋盖通过锚定 crRNA:靶链双链并引导靶链向 RuvC 核心移动,在 DNA 靶链的移动中起着关键作用,DNA 切割实验也证实了这一点。在这种机制中,REC2 结构域将 crRNA:靶链双链推向酶的核心,而 Nuc 结构域通过向内弯曲来帮助靶链在 RuvC 核心内的弯曲和调节。了解 Cas12a 活性背后的这一关键过程将丰富基础知识并促进进一步的基因组编辑工程策略。

恶性疟原虫 DNA/RNA 异位过度表达
摘要 含有 Alba 结构域的蛋白质在古细菌和真核生物中普遍存在。通过与 DNA、RNA 或 DNA:RNA 杂交体结合,这些蛋白质在基因组稳定、染色质组织、基因调控和/或翻译调节中发挥作用。在疟原虫恶性疟原虫中,已描述了六种 Alba 结构域蛋白 PfAlba1–6,其中 PfAlba1 已成为
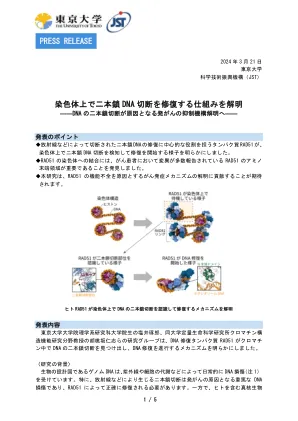
了解修复染色体上双链DNA断裂的机制
(注2)核小体这是染色质的基本单位,是一种结构,其中大约150个DNA碱基对包裹在一个组蛋白八聚体周围,该组蛋白八聚体包含两个分子(H2A,H2B,H2B,H3,H4)中的四种分子。 (注3)冷冻电子显微镜A显微镜,其中包含蛋白质样品在极端低温的环境中冷冻,并用电子束观察到限制样品。通过拍摄大量图像,可以获得具有多种角度信息的粒子图像,并且可以从该信息中重建样品的三维结构。 (注4)氨基末端结构域(N末端结构域)在蛋白质末端的一个区域,该区域具有氨基群,最初是在蛋白质合成过程中合成的。 RAD51由两个球状结构域组成,其中一个球状结构域存在于氨基末端,一个与RECA同源的球状结构域。 (注5)L1回路区域该区域在与RECA同源的球状结构域中发现,对于与线性DNA结合很重要。联系(请联系演讲者以获取研究详细信息)Kurumizaka hitoshi教授,定量生命科学研究所,东京大学电话:03-5841-7826传真:03-5841-1468电子邮件:kurumizaka:kurumizaka [at] iqb.u-tokyo.ac.ac.jp procention nocation nocation jst Impaction jst Impact项目> Fumie Imabayashi电话:03-3512-3528传真:03-3222-2068电子邮件:Eratowww [at] jst.go..jp <与报告相关的询问>通用事务团队,定量生命科学研究所,东京大学电话:03-5841-781-781-781313 soumu [at] iqb.u-tokyo.ac.ac.jp日本科学技术局公共关系部电话:03-5214-8404传真:03-5214-8432电子邮件:

通过改变LAZY1中的一个氨基酸来抑制水稻地上部分向地性
摘要:分蘖角度是决定禾谷类作物株型和产量的重要性状。在重力刺激下,分蘖角度部分由LAZY1(LA1)蛋白在细胞核和质膜之间的动态重新分配来控制,但其潜在机制尚不清楚。在本研究中,我们基于对水稻(Oryza sativa L.)扩散分蘖突变体la1 G74V的分析,鉴定并描述了LA1的一个新的等位基因,该突变体在该基因预测的跨膜(TM)结构域编码区中发生非同义突变。该突变导致地上部重力性完全丧失,从而导致植物匍匐生长。我们的研究结果表明,LA1不仅定位于细胞核和质膜,而且定位于内质网。去除LA1中的TM结构域会使植物表现出与la1 G74V相似的扩散分蘖表型,但不影响质膜定位;因此,它与玉米中的直系同源物 ZmLA1 有区别。因此,我们认为 TM 结构域对于 LA1 的生物学功能是必不可少的,但该结构域并不决定蛋白质在质膜上的定位。我们的研究为 LA1 介导的地上性调控提供了新的见解。

对“无药可用”的致病蛋白质进行药物治疗:重点...
信号转导和转录激活因子 (STAT) 3 被归类为“不可用药”的致病蛋白,尽管它含有 Src 同源性 (SH) 2 结构域,这是一个潜在的致命弱点,过去 30 年来一直未能被学术界和制药界成功针对。基于突变和建模研究,我们的小组开发了一种独特的虚拟配体筛选策略,针对 STAT3 SH2 结构域,结合强大的生化和细胞分析以及基于结构的药物化学,并鉴定出 TTI-101。TTI-101 是临床开发中最先进的直接小分子含 SH2 结构域致病蛋白抑制剂之一。TTI-101 目前正在进行 1 期研究,以确定其在晚期实体瘤患者中的安全性和耐受性以及药效学作用和疗效。