XiaoMi-AI文件搜索系统
World File Search System肌动蛋白

慢病毒介导的 CRISPR/Cas9 递送可降低小鼠青光眼模型中的眼压
肌动蛋白 ( MYOC ) 突变是已知的原发性开角型青光眼的主要遗传原因,约占所有病例的 4%。MYOC 突变会导致功能获得性表型,其中突变的肌动蛋白会在内质网 (ER) 中积聚,导致 ER 应激和小梁网 (TM) 细胞死亡。因此,在基因组水平上敲除肌动蛋白是永久治愈该疾病的理想策略。我们之前已成功利用 CRISPR/Cas9 基因组编辑通过腺病毒 5 (Ad5) 靶向 MYOC。但是,Ad5 不是适合临床使用的载体。在这里,我们试图确定腺相关病毒 (AAV) 和慢病毒 (LV) 靶向 TM 的功效。首先,我们通过玻璃体内 (IVT) 和前房 (IC) 注射检查了单链 (ss) 和自互补 (sc) AAV 血清型以及表达 GFP 的 LV 的 TM 趋向性。我们观察到 LV_ GFP 表达对通过 IVT 途径注射的 TM 更具特异性。色氨酸突变体 scAAV2 的 IC 注射显示 TM 中 GFP 的显著表达。然而,在睫状体和视网膜中也观察到了强劲的 GFP 表达。我们接下来构建了表达 Cas9 和靶向 MYOC (crMYOC) 的引导 RNA (gRNA) 的慢病毒颗粒,并用 LV_cr MYOC 转导稳定表达突变型肌动蛋白的 TM 细胞可显著减少肌动蛋白积累及其相关的慢性 ER 应激。在 Tg-MYOC Y437H 小鼠中单次 IVT 注射 LV_cr MYOC 可减少 TM 中的肌动蛋白积累并显著降低升高的眼压。总之,我们的数据表明,LV_cr MYOC 靶向 TM 中的 MYOC 基因编辑并挽救了肌动蛋白相关青光眼的小鼠模型。

CAPG 是埃博拉病毒感染所必需的,它通过控制病毒从受感染细胞中流出
摘要:埃博拉病毒 (EBOV) 的复制依赖于肌动蛋白的功能,尤其是在通过巨胞饮作用进入细胞和病毒从细胞中释放时。此前,参与肌动蛋白成核的主要肌动蛋白调节因子,如 Rac1 和 Arp2/3,在这两个步骤中都发挥着重要作用。然而,在成核的下游,需要许多其他细胞因子来控制肌动蛋白动力学。这些如何调节 EBOV 感染仍不清楚。在这里,我们确定了肌动蛋白调节蛋白 CAPG 对 EBOV 复制很重要。值得注意的是,敲低 CAPG 特异性地抑制了病毒的传染性和感染性颗粒的产量。基于细胞的机制分析表明,从受感染的细胞中产生病毒需要 CAPG。邻近连接和裂解绿色荧光蛋白重建试验表明,CAPG 与 VP40 紧密结合,这种结合是通过 CAPG 的 S1 结构域介导的。总体而言,CAPG 是一种新型宿主因子,通过将肌动蛋白丝稳定性与病毒从细胞中排出相连接来调节 EBOV 感染。

肌动蛋白细胞骨架在蝴蝶翼尺度中的结构颜色形成中扮演多个角色
黄油中的生动结构颜色是由光子纳米结构散射光引起的。结构颜色用于众多生物信号功能,并具有重要的技术应用。从光学上讲,这种结构是充分理解的,但是对它们在体内发展的洞察力仍然很少。我们表明,肌动蛋白与黄油翼鳞片中的结构颜色形成密切相关。使用成人和发展中H. sara的虹彩(结构上有色)和非冰箱尺度之间的比较,我们表明虹彩尺度具有更密集的肌动蛋白束,导致倾斜脊密度增加。超分辨率的微分析跨三个遥远相关的黄油种类揭示,肌动蛋白在尺度发育过程中反复重新安排,并且在形成光学纳米结构时至关重要。此外,在这些后期的发育阶段进行肌动蛋白扰动实验导致H. Sara的结构颜色几乎几乎总损失。总体而言,这表明肌动蛋白在黄油含量尺度的结构颜色形成过程中起着至关重要的直接模板作用,从而提供了在鳞翅目中可能具有普遍性的脊模式机制。

线粒体MICOS复合基因,涉及左心脏综合征,保持心脏收缩和肌动蛋白完整性
1开发,老化和再生计划,遗传疾病与衰老研究中心,桑福德·伯纳姆·普雷比斯医学发现研究所,美国圣地亚哥; 2美国圣地亚哥医学院桑福德再生医学联盟生物工程系; 3美国罗切斯特梅奥诊所的心血管遗传学研究实验室; 4美国罗切斯特梅奥诊所定量健康科学系计算生物学系; 5美国圣地亚哥的拉迪医院MC 5004儿科医学院儿科医学院; 6美国罗切斯特市梅奥诊所分子和药理学系儿科和青少年医学系儿科心脏病学再生医学中心,分子与药理学系和实验治疗师; 7儿科和青少年医学系心血管医学系,心血管遗传学研究实验室,美国罗切斯特梅奥诊所

靶向肌球蛋白结合蛋白C
抽象心脏MYBP-C(CMYBP-C)与肌动蛋白 - 肌球蛋白相互作用,以微调心脏肌肉收缩。CMYBP-C的磷酸化降低了CMYBP-C与肌动蛋白或肌球蛋白的结合,通常会减少心力衰竭(HF)患者,并且在HF模型系统中受到心脏保护。因此,CMYBP-C是模仿磷酸化和/或扰动其与肌动蛋白或肌球蛋白相互作用的HF药物的潜在靶标。我们用荧光素-5-甲酰亚胺(FMAL)和CMYBP-C(CC0-C2)的C0-C2片段标记了肌动蛋白(CC0-C2)。我们在FDA批准的药物库上进行了两个互补的高通量筛选(HTS),以发现专门结合CMYBP-C的小分子,并使用荧光寿命(FLT)检测来影响其与肌动蛋白或肌球蛋白的相互作用。我们首先激发了FMAL并检测到其FLT,以测量荧光共振能量转移(FRET)的变化,从FMAL(供体)到TMR(受体),表明蛋白质复合物中的结合和/或结构变化。使用相同的样品,然后使用更长的波长激光直接激发TMR,以检测化合物对TMR环境敏感FLT的影响,以识别直接结合CC0-C2的化合物。次要测定,对主要HTS分析的选定调节剂进行,这些化合物的特异性是磷酸化对未磷酸化的CC0-C2的特异性,而CC0-C2对C1-C2与C1-C2相对于快速骨骼肌的C1-C2(FSKC1-C2)。确定的化合物的子集调制了心脏和/或骨骼肌原纤维中ATPase活性。简介这些测定法确定了发现CMYBP-C-肌动蛋白/肌球蛋白相互作用的小分子调节剂的可行性,其最终目的是开发HF疗法。
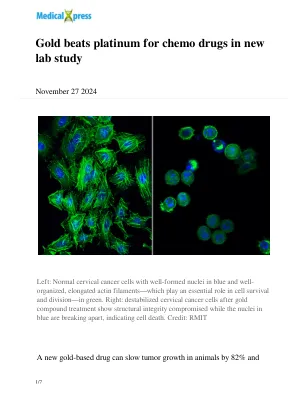
新实验室研究表明黄金在化疗药物中的应用优于铂金
左图:正常宫颈癌细胞,细胞核结构良好(蓝色),肌动蛋白细丝(绿色)排列整齐,肌动蛋白细丝在细胞存活和分裂中起着至关重要的作用。右图:不稳定的宫颈癌细胞经金化合物处理后,结构完整性受损,细胞核(蓝色)分裂,表明细胞死亡。图片来源:RMIT

CRACD损失诱导肺腺癌的神经内分泌细胞可塑性
先前,我们鉴定了CRACD(抑制肌动蛋白动力学调节剂,也称为Crad/Kiaa1211)肿瘤抑制剂,该肿瘤抑制剂,该抑制剂通过结合和抑制限制蛋白来促进肌动蛋白聚合以促进肌动蛋白以促进肌动蛋白聚合23。有趣的是,我们观察到CRACD KO小鼠肺中的增生病变23。这种观察结果使我们假设CRACD损失可能会驱动肺中的NE样细胞可塑性。为了测试这一点,我们检查了CRACD KO小鼠肺组织。与CRACD野生型(WT)不同,CRACD KO肺组织在支气管呼吸道和肺泡中表现出NE样增生(图1a)。免疫荧光(IF)染色证实了这种NE样细胞质量的增殖性质,如MKI67+所示,MKI67+是细胞增殖的标志物。此外,质量表达了几个NE标记,包括KRT19,SYP,CGRP,CHGA和ASCL1(图。

太赫兹辐射能量通过水层传播:破坏活细胞中的肌动蛋白丝
近几十年来,随着太赫兹 (THz) 光源的发展,工业和医学应用相继被提出。此外,THz 辐射对人体健康的毒性也引起了在此频率区域工作的研究人员的浓厚兴趣 1 。两个项目,欧洲 THz-BRIDGE 和 SCENIHR 的国际 EMF 项目 2 ,总结了近期有关 THz 辐射对人体影响的研究。例如,THz 波对 DNA 稳定性产生非热影响 3 – 5,这可能导致人类淋巴细胞的染色体畸变 6 。还证明了小鼠皮肤中伤口反应基因的转录激活 7 和人造人体 3D 皮肤组织模型 8 中的 DNA 损伤。大多数研究集中在上皮和角膜细胞系,因为在这个频率区域液态水的强烈吸收下,THz 光子在组织表面被完全吸收。但是,如果将 THz 辐射转换为可以传播到水中的另一种能量流,THz 波的照射可能会对组织内部造成损伤。事实上,THz 光子能量一旦被体表吸收,就会转换为热能和机械能。我们最近观察到 THz 脉冲在液态水表面产生冲击波 9 。产生的冲击波可以传播几毫米深。类似的现象也可能发生在人体上。THz 诱导的冲击波会对生物分子产生机械应力并改变其形态。THz 辐射的这种间接影响尚未被研究过。为了揭示 THz 诱导的冲击波对生物分子的影响,我们重点研究了肌动蛋白的形态。肌动蛋白有两种功能形式,单体球状 (G)-肌动蛋白和聚合丝状 (F)-肌动蛋白。肌动蛋白丝形成复杂的细胞骨架网络,在细胞形状、运动和分裂中起着至关重要的作用 10 。使用肌动蛋白的一个优点是,我们可以很容易地从组织中获得足够的纯化 G- 肌动蛋白 11 ,以重建体外聚合反应。肌动蛋白丝可以通过用硅-罗丹明 (SiR)-肌动蛋白染色直接在荧光显微镜下观察 12 。由于肌动蛋白在正常和病理细胞功能中起着关键作用,包括转录调控、DNA 修复、癌细胞转移和基因重编程 13 - 16 ,各种化合物和调节蛋白已被分析用于研究和治疗目的 17 。在这项研究中,我们调查了 THz 诱导的冲击波对肌动蛋白丝的影响

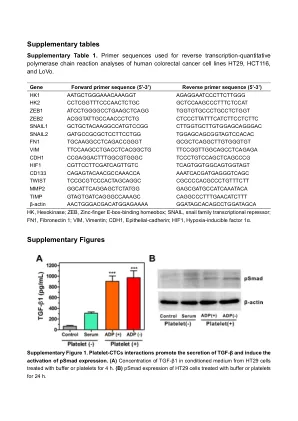
补充表
ctctttctctctc nail1 gctgctgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtggtggggggghts snal2 TGCAAGCCTCGGGGTGTGTGGTGGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGCCCCCCCCCCCCCGCTGGTGGCTGGTGTGTGTGTGGTGH1 CDH1 CDGGTGGGGC TCCTGTCCAGCCCCGCCGCCCGCGCGCGCCGCGGS CGTTCTCTCTGTGTC TCAGTGGGGGGGGGTGGTGTGTGTGTs CD133 CAGTACCAACCAAACCIGITCIGITITIs GAGCIGITING TIMP GTAGGGGGGGGGGGGGGGGGGCAGC caggccttttgttttttttttttttttttttttttsβ-肌动蛋白actghtgacity gtatagcagcatgityctctttctctctc nail1 gctgctgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtgtggtggggggghts snal2 TGCAAGCCTCGGGGTGTGTGGTGGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGTGCCCCCCCCCCCCCGCTGGTGGCTGGTGTGTGTGTGGTGH1 CDH1 CDGGTGGGGC TCCTGTCCAGCCCCGCCGCCCGCGCGCGCCGCGGS CGTTCTCTCTGTGTC TCAGTGGGGGGGGGTGGTGTGTGTGTs CD133 CAGTACCAACCAAACCIGITCIGITITIs GAGCIGITING TIMP GTAGGGGGGGGGGGGGGGGGGCAGC caggccttttgttttttttttttttttttttttttsβ-肌动蛋白actghtgacity gtatagcagcatgity