XiaoMi-AI文件搜索系统
World File Search System行进

指数尾部的组合行进
观察到儿童化学套装中的成分可以创造出比宇宙中原子更多的不同组合。基于这一见解,Weitzman (1998) 构建了一个增长模型,其中新想法是旧想法的组合。然而,由于组合增长如此之快,他发现增长受到我们处理爆炸式增长的想法数量的限制,而组合学在确定增长率方面基本上没有发挥任何正式作用:有如此多的潜在组合,以至于数量不是限制因素。组合过程没有发挥更核心的作用,这有点令人失望和困惑。另一篇文献强调了指数增长和帕累托分布之间的联系。具体来说,Kortum (1997) 引入了一种建模经济增长的新方法,并认为帕累托分布至关重要:如果生产率是在从某个分布中抽取的多个样本中取的最大值(只使用最好的想法),那么在他的设置中,生产率的指数增长要求抽取的次数呈指数增长,并且所抽取的分布是帕累托分布,至少在上尾是这样。有趣的是,似乎需要如此强的分布假设。也许提取想法的底层分布是帕累托分布,但为什么会这样呢?毕竟,在经济学的许多其他应用中,帕累托分布是推导出来的,而不是假设的。例如,Gabaix (1999)、Luttmer (2007) 以及 Jones 和 Kim (2018) 强调,城市规模、公司就业、收入和财富都具有帕累托分布的特征。但是,该文献显示了这些帕累托分布是如何作为内生结果出现的。这就引发了一个问题:帕累托分布在 Kortum 方法中是否真的是必要的。而且,Romer 和 Weitzman 认为组合学应该是理解增长的核心,那么他们的观点又怎么了?本文结合 Kortum (1997) 和 Weitzman (1998) 的观点来回答这些问题。假设创意是现有成分的组合,就像菜谱一样。每个菜谱的生产率都是从概率分布中得出的。与 Romer 和 Weitzman 的观点一样,我们可以从现有成分中创造出的组合数量大到本质上是无限的,而我们受限于处理这些组合的能力。令 N t 表示截至日期 t 已经评估过的菜谱成分数量。换句话说,我们的“食谱”包括了所有可能由 N t 种原料组成的食谱:如果每种原料都可以加入或排除在食谱之外,那么食谱中总共有 2 N t 种食谱。最后,研究包括将新食谱添加到食谱中,即评估它们并了解它们的生产力。特别是,假设研究人员在食谱中添加新配料,并了解其生产率,使得 N t 呈指数增长。我们称一个包含 2 N t 个食谱的设置

使用快速行进方法
抽象有效且智能的路径规划算法设计用于在动态海洋环境中进行操作,对于无人体表面车辆(USV)的安全操作至关重要。当前的大多数研究都通过基于解决方案为基于每个USV都有强大的通信渠道以获取基本信息(例如海上车辆的位置和速度)的非执行假设来涉及“动态问题”。在本文中,提出了基于卡尔曼过滤器的预测路径计划算法。该算法旨在预测移动船的轨迹以及实时的USV自身位置,并因此评估碰撞风险。对于计划计划的路径,提出并开发了一种加权快速的正方形方法,以搜索最佳路径。可以通过调整加权参数来针对任务要求(例如最小旅行距离和最安全路径)进行优化路径。已使用包括实际环境方面的许多模拟对所提出的算法进行了验证。结果表明,算法可以充分处理复杂的交通环境,并且生成的实用路径适合于无人驾驶和载人船只。
高阶空间行进算法的改进...
美国宇航局的低轰飞行演示 (LBFD) 项目 o 主要目标是演示在降低响度水平下进行超音速陆上飞行的可行性 o X-59 静音超音速技术 (QueSST) 飞机 o 任务规划需要包含 O (1000)- O (10,000) 个解决方案的大型数据库 • 远离机身的细网格以跟踪冲击 • 高计算资源 • 必须准确 • 必须自动化

dmt改变皮层行进波
抽象的许多生物在自然界中具有进化的机制,可以耐受严重的低氧或缺血,包括具有冬眠能力的北极松鼠(AGS)。尽管AGS中的低氧或缺血耐受性涉及生理适应性,但对内在AGS细胞弹性对代谢应激的弹性的关键细胞机制知之甚少。通过基于细胞存活的cDNA表达筛选在神经祖细胞中,我们确定了AGS ATP5G1的遗传变异,该变异赋予了细胞对代谢应激的弹性。ATP5G1编码线粒体ATP合酶的亚基。在小鼠细胞中的异位表达和内源性AGS基因座的CRISPR/CAS9基础编辑表明,一个AGS特异性氨基酸取代在介导AGS ATP5G1的细胞保护过程中的因果作用。AGS ATP5G1通过调节线粒体形态变化和代谢功能来促进代谢应激的弹性。我们的结果从哺乳动物的冬眠器中确定了ATP5G1的自然发生的变体,该变体对代谢应激有助于固有的细胞保护作用。

通过分析方法和行进优化 GEO 带观测
观测和编目 GEO 带中的卫星对于空间卫星跟踪、避免碰撞和空间态势感知应用来说是一项至关重要的工作。然而,由于卫星数量庞大且需要精确的观测规划,因此对 GEO 区域内的卫星进行有效和全面的观测带来了巨大的后勤挑战。传统的卫星观测方法通常涉及静态望远镜定位或次优手动扫描技术,这些技术可能既耗时又低效。此外,鉴于卫星轨道的动态性质和 GEO 带中卫星的庞大数量,需要采用创新方法来优化观测策略。为了应对这些挑战,通过使用分析方法减少夜空中观测的位置数量来优化观测,为在 GEO 带内编目卫星提供了一种系统的方法。通过将旅行商问题 (TSP) 的原理与分析方法和望远镜技术相结合,我们旨在最大限度地缩短观测目标之间的过渡时间或摆动,同时最大限度地提高沿优化观测路径的数据采集效率。

改进行进立方体算法的修改规则代表3D
摘要:行进立方体是3D重建的最广泛使用的等曲面算法。在案例研究中,本文使用了来自大脑图像的MRI的医学数据,尤其是在call体(CC)部分中,以及来自Stagbeetle数据集的音量数据。选择此案例研究以突出3D图像可视化的临床重要性。这项研究可以通过显示固体解剖形状和位置来帮助,这可以指导脑损伤的位置,而小于1 mm的较小误差;因此,它可以支持和最大程度地减少脑外科手术的风险。案例研究是称为call体的大脑的一部分,通常用作脑部手术的参考。对于输入数据,本文使用深度学习方法使用2D分割来获得CC段。本文使用120名患者,培训80%,在国家医院进行测试20%。本文发现了11个矢状切片,其中包含每位患者的166个切片中的call体。这项工作提出了一种改进的MC算法,为现有规则增加了20个新规则,加强了Voxel代表的规则,并将原始的Martinging Cubes算法的15条规则增加到35。因此,3D重建模型覆盖了大孔,使其在很大程度上固体。拟议的3D可视化实现了来自国家医院的数据集的零开放边缘。结果表明,应用改进的行进立方体算法产生了一个3D表示,其结果更好,更健壮,这证明了存在更多的顶点和三角形以及不存在开放边缘的情况。高级游行立方体是拆除开放边缘的好方法。
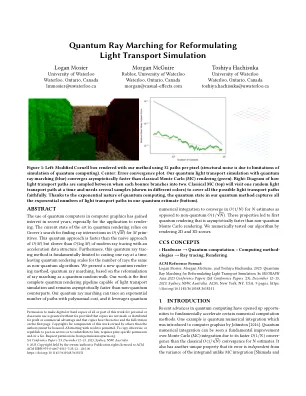
量子射线行进以重新设计光传输模拟
图1:左:使用我们的方法呈现的修改后的康奈尔盒,使用每个像素的32个路径(结构噪声是由于量子计算模拟的局限性引起的)。中心:错误收敛图。我们使用量子射线行进(蓝色)的量子光传输模拟比古典蒙特卡洛(MC)渲染(绿色)快地收敛。右:在每个弹跳分支分为两者之间如何在两者之间采样光传输路径的图。古典MC(顶部)将一次访问一个随机的光传输路径,需要几个样本(以不同的颜色显示),以忠实涵盖所有可能的轻型运输路径。由于量子计算的指数性质,我们的量子方法中的量子状态在一个量子估计中捕获了所有指数的光传输路径(底部)。

基于快速行进方法和深度强化学习的自主船的新颖道路遵循方法
路径以下是自动船只的必不可少的工具之一,它确保自动船能够充分能够在指定的无碰撞水中导航。这项研究提出了一种新的路径,遵循基于快速行进方法(FM)方法和深入增强学习(DRL)的自主船的方法。所提出的方法能够控制船以遵循不同的路径,并确保路径跟踪误差始终在设定范围内。借助FM方法,基于网格的路径偏差图是专门生成的,以指示网格点和路径之间的最小差异。此外,特定设计的路径偏差感知器是为了模拟基于路径偏差图的设定路径偏差边界的范围传感器。之后,培训了一个基于DRL的圆形路径来控制船舶。尤其是通过模拟对该方法进行验证和评估。获得的结果表明,所提出的方法始终能够维持较高的总体效率,并具有相同的层次,以遵循不同的路径。此外,这种方法的能力对自主船的发展表现出显着的贡献。

人的大脑在每个时间的瞬间通过行进波的多路复用编码视觉事件的编年史
人脑连续处理视觉输入的流。然而,单个图像通常会触发延伸超过1s的神经反应。要了解大脑如何编码和保持连续的图像,我们用脑电图分析了人类受试者观看时的大脑活动。 5000个视觉刺激以快速序列呈现。首先,我们确认可以从大脑活动中解码每种刺激; 1s,我们证明大脑在每次瞬间同时代表多个图像。第二,我们在预期的视觉层次结构中进行了定位的脑反应,并表明在每次瞬间,不同的大脑区域代表了过去刺激的不同快照。第三,我们提出了一个简单的框架,以进一步表征这些行进波的动态系统。我们的结果表明,一系列神经回路,每个链由(1)隐藏的维护机制和(2)可观察到的更新机制组成,它解释了视觉序列引起的宏观脑表示的动力学。一起,这些结果详细介绍了一个简单的体系结构,解释了如何同时在大脑中同时代表连续的视觉事件及其各自的时间。

2024-50-Z Union Woodlands PUD 保护区
从上述东北区东北角的一块石头开始,沿区段线向南向东 0 度 01 分 18 秒(假定方位)行进 1982.20 英尺到起点,从所述起点继续沿区段线向南向东 0 度 01 分 18 秒行进 656.11 英尺;然后沿四分之一区段线向南向西 89 度 08 分 28 秒行进 2263.97 英尺(实际距离)(先前契约为 2263.72 英尺);然后沿美国 421 号公路中心线向北向西 20 度 31 分 22 秒行进 1082.86 英尺(实际距离)(先前契约为 1082.04 英尺);然后向北向东 89 度 08 分 28 秒行进 1297.50 英尺;然后向南向西 0 度 06 分 52 秒行驶 10.57 英尺;再向南向东 89 度 16 分 46 秒行驶 683.49 英尺;再向南向西 0 度 46 分 39 秒行驶 344.18 英尺;再向北向东 88 度 27 分 47 秒行驶 667.93 英尺到达起点,面积大约为 51.5957 英亩。