XiaoMi-AI文件搜索系统
World File Search System运动神经元
神经元
继续需要动物来提高大脑研究,朱迪思·霍姆伯格1*,罗杰·阿丹7,纳塔利亚·阿伦娜15,安东尼斯·阿西米纳斯21,迈克尔·巴德15,汤姆·贝克斯18,迪诺瓦·P·贝格·P·贝吉·贝吉·布洛克兰,阿尔扬·布洛克兰10,E.Gert,E.洛斯·菲茨(Los P. 1,Sharon M. Kolk 2,Aniko Korosi,13,Meziel Mechicz。 ,Umberto Olcese 5,Azahara Oliva 32,Jocelien Olivier 6,Massimo Pasqualetti 23,Cyriel Ma Pennartz 5,Piotr Popik 25,Jos Prickaerts 10,Liset M de La la la Prida Prida Prida Prida 29 MJ Vanderschuren 11,Tomonori Takeuchi 22,Rixt van der Veen 5,Marten P. Smidt 5,Vladys Vyavski 19,Maximi Coryski J. Wierenga 11,Bella Williams 20,Ingo Willuhn,Ingo Willuhn 8,9,MarkusWöhr13,14,14,14,21,Monique Woldevel * van * van * van n van * zel@donders.ru.nl,朱迪思。

无机砷改变多巴胺能神经元的发育,但不改变血清素能神经元的发育,并通过 Sonic Hedgehog 通路诱导斑马鱼的运动神经元发育
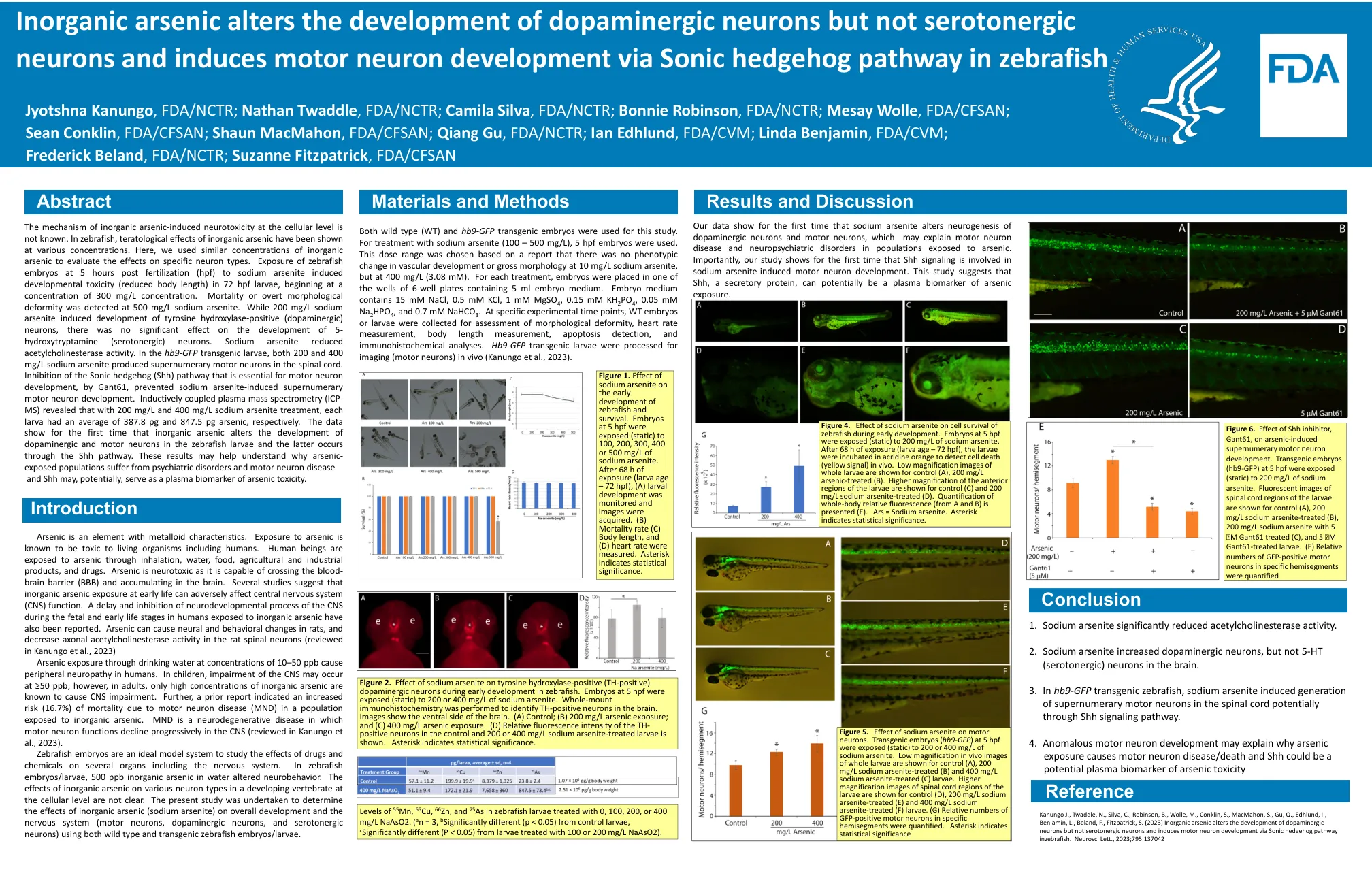
无机砷在细胞水平上诱发神经毒性的机制尚不清楚。在斑马鱼中,不同浓度的无机砷均有致畸作用。在这里,我们使用了类似浓度的无机砷来评估其对特定神经元类型的影响。受精后 5 小时 (hpf) 的斑马鱼胚胎暴露于亚砷酸钠中,在 72 hpf 幼虫中诱发发育毒性(体长缩短),浓度从 300 mg/L 开始。在 500 mg/L 亚砷酸钠下检测到死亡或明显的形态畸形。虽然 200 mg/L 亚砷酸钠诱导酪氨酸羟化酶阳性(多巴胺能)神经元的发育,但对 5-羟色胺(血清素能)神经元的发育没有显著影响。亚砷酸钠降低了乙酰胆碱酯酶活性。在hb9-GFP转基因幼鱼中,200和400mg/L亚砷酸钠均在脊髓中产生了多余的运动神经元。通过Gant61抑制运动神经元发育所必需的Sonic Hedgehog(Shh)通路,可以阻止亚砷酸钠诱导的多余运动神经元发育。电感耦合等离子体质谱(ICP-MS)分析表明,在200mg/L和400mg/L亚砷酸钠处理下,每只幼鱼平均砷含量分别为387.8pg和847.5pg。数据首次表明无机砷改变斑马鱼幼鱼多巴胺能神经元和运动神经元的发育,后者是通过Shh通路发生的。这些结果可能有助于理解为什么接触砷的人群会患上精神疾病和运动神经元疾病,并且 Shh 可能潜在地充当砷毒性的血浆生物标志物。

脑损伤后运动前皮质神经元活动的快速和双半球重组
脑损伤会导致远离病变的几个区域发生血流动力学变化。我们的目标是更好地了解清醒且行为正常的雌性猴子中这种重组的神经元相关性。我们使用可逆失活技术“损伤”初级运动皮层,同时在行为障碍发生之前和之后连续记录两个半球腹侧运动前皮层的神经元活动。失活迅速引起神经元放电的深刻改变,这些改变在每个半球内和两个半球之间都是不均匀的,发生在受影响或未受影响的手臂运动期间,并在抓握的不同阶段有所不同。我们的研究结果支持了广泛的、比预期更复杂的神经元重组发生在双半球皮层网络中参与控制手部运动的保留区域中。这种广泛的重组模式提供了潜在的目标,应该考虑开发脑损伤后早期应用的神经调节方案。

人类运动单位放电模式揭示了跨收缩水平向运动神经元的神经调节和抑制驱动的差异
所有电动机命令都会收敛到电动机单元(MUS),这些电动机将信号转移到肌肉纤维的机械作用中。由于离子(兴奋性/抑制性)和代谢性(神经调节)输入的组合,此过程是高度非线性的。神经调节输入有助于树突持续的内向电流,这引入了MU放电模式中的非线性,并为运动命令的结构提供了见解。在这里,我们研究了神经调节的相对贡献和抑制模式,以最大70%的收缩力调节人MU排出模式。利用从三种人体肌肉(胫骨前 - TA,ta和巨大的外侧和内膜)鉴定出的MU排出模式,我们表明,随着收缩力增加,发作偏移率滞后率(ΔF)增加了升级的MU放电模式,而s升则增加了线性,并较低。在后续实验中,我们证明了增加δF的观察结果和更线性的上升MU放电模式,即使在收缩持续时间和力率增加时,也可以保持更大的收缩力。然后,我们使用在硅运动神经池中高度逼真的逆转ta Mu放电模式来证实人类记录中推断出的生理机制。我们证明了一个严格的限制性解决方案空间,通过这种空间,只有通过增加的神经调节和更相互的互惠来重新创建收缩力引起的实验获得的MU放电模式的变化(即推扣)抑制模式。总而言之,我们的实验和计算数据表明,神经调节和抑制模式的形状是独特的,以产生放电模式,这些模式支持力在大部分运动池募集范围内增加。

- 运动神经元疾病的口腔吸力 组织捐赠 - 信息表I 加速运动神经元治疗的研究... 英国MND研究所 倒计时 MND协会关于使用干细胞的政策声明,如果我们要停止运动神经元疾病
在人无法同意但不执行程序会损害其健康状况的情况下,该人的专家团队将建议吸力是否合适。如果该人没有能力做出决定,则需要根据《心理能力法》的最大利益决定,除非是紧急情况。必须检查一个人是否记录了拒绝治疗(ADRT)或紧急医疗计划的预先决定,以及已包括了哪些治疗或干预措施。

升高的NLRP3炎性体激活与ALS中的运动神经元变性有关
1鲁尔 - 大学Bochum解剖研究所细胞学系,德国Bochum 44801; hilalcihankaya@gmail.com(H.C.); carsten.theiss@rub.de(C.T。)2国际神经科学研究生院(IGSN),Ruhr-University Bochum,44801 Bochum,德国; konstanze.winklhofer@rub.de 3分子细胞生物学系,生物化学与病原体化学研究所,医学院,Ruhr-University Bochum,44801 Bochum,Bochum,德国; verian.bader@rub.de 4神经病学系,海默尔肌肉研究所,大学医院,伯格曼·史密尔,鲁尔 - 大学Bochum,Buerkle-de-de-la-camp-platz-1,44789 Bochum,Bochum bochum; matthias.vorgerd@bergmannsheil.de 5细胞生物学研究所(癌症研究),大学医院埃森大学,杜伊斯堡 - 埃森大学,德国45147,德国埃森; johann.matschke@uk-esen.de *通信:veronika.matschke@rub.de;电话。: +49-234-32-25018

通过终端选择器功能中的时间模块化运动神经元身份的建立和维护
摘要在胃肠道中,神经rest细胞在神经板边界处指定为PAX7表达。使用单细胞RNA测序与高分辨率原位杂交结合以识别新型的转录调节剂,我们表明染色质重塑剂HMGA1在规格之前高度表达并保持在迁移的鸡神经trest细胞中。暂时控制的CRISPR-CAS9介导的敲除在神经Crest发育中发现了HMGA1的两个不同功能。在神经板边界,HMGA1调节依赖PAX7的神经rest谱系规范。在移民阶段,第二个角色表现出HMGA1损失减少了独立于PAX7的背神经管的颅顶移民。有趣的是,这是通过稳定的ß-catenin挽救的,因此将HMGA1作为规范WNT激活剂。一起,我们的结果表明,HMGA1在神经Crest发育过程中以双峰方式起作用,以调节神经板边界的规范,然后通过规范WNT信号传导从神经管中移民。

ERA-NET神经元
国家卫生和医学研究委员会 (NHMRC) 澳大利亚 科学研究基金会 (FRS-FNRS) 比利时 科学、教育和青年部 (MSEY) 克罗地亚 法国国家研究机构 (ANR) 法国联邦教育和研究部 (BMBF) 德国 德国研究基金会 (DFG) 德国国家研究、发展和创新办公室 (NKFIH) 匈牙利 卫生研究委员会 (HRB) 爱尔兰 卫生部首席科学家办公室 (CSO-MOH) 以色列 意大利卫生部 (IT-MoH) 意大利 拉脱维亚科学理事会 (LZP) 拉脱维亚 立陶宛研究理事会 (LMT) 立陶宛 挪威研究理事会 (RCN) 挪威国家研究与发展中心 (NCBR) 波兰* 高等教育、研究、发展和创新资助执行机构 (UEFISCDI) 罗马尼亚

初级运动皮层锥体束神经元细胞兴奋性降低,导致模拟帕金森病运动皮层网络活动反常增加
。CC-BY 4.0 国际许可证可在未经同行评审认证的情况下获得)是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是