XiaoMi-AI文件搜索系统
World File Search System骨细胞

骨头
脑衍生的神经营养因子(BDNF)是一种神经营养蛋白,在中枢神经系统和周围组织中表达,受到GSα /CAMP途径的调节。在骨骼中,它调节成骨,并刺激骨化肿瘤(如多发性骨髓瘤)中的RANKL分泌和破骨细胞形成。纤维发育不良(FD)是由GSα基因的功能收益突变引起的罕见的骨骼遗传疾病,其中RANKL依赖性增强的骨吸收是骨骼脆弱性和临床发病率的主要原因。我们观察到BDNF转录本在人类FD病变中表达。具体而言,对从FD患者获得的活检进行的免疫定位研究揭示了成骨细胞中BDNF的表达,并且在纤维组织内的纺锤形细胞中的表现较低。因此,我们假设BDNF可以通过刺激RANKL分泌和骨吸收来在FD的发病机理中发挥作用。为了测试这种疗法,我们使用了人类疾病的EF1α-GSαR201C小鼠模型(FD小鼠)。Western印迹分析显示,与WT小鼠相比,FD小鼠的骨段中BDNF的表达更高,而小鼠FD病变中的免疫标记模式与在人FD中观察到的相似。用抗BDNF的单克隆抗体对FD小鼠进行处理,可减少纤维组织,以及股骨病变内的破骨细胞和骨爆炸的数量。这些结果揭示了BDNF是FD发病机理的新玩家,并且可以在FD骨骼病变中滋养破骨细胞生成的潜在分子机制。他们还建议BDNF抑制作用可能是减少FD中异常骨骼重塑的一种新方法。

自噬和骨骼疾病
références1。Mizushima N,Levine B,Cuervo AM,Klionsky DJ。自噬通过细胞自我消化与疾病作斗争。自然。2008年2月28日; 451(7182):1069–75。 2。 Mizushima N,Komatsu M.自噬:细胞和组织的翻新。 单元格。 2011年11月11日; 147(4):728–41。 3。 Pierrefite-Carle V,Santucci-Darmanin S,Breuil V,Camuzard O,Carle GF。 骨骼中的自噬:保持平衡。 老化Res Rev. 2015年11月; 24(pt b):206-17。 4。 Liu F,Fang F,Yuan H等。 通过抑制成骨细胞末端分化,FIP200缺失对自噬的抑制导致小鼠的骨质减少。 J骨矿工销售J Am Soc Bone Miner Res。 2013年11月; 28(11):2414–30。 5。 Nollet M,Santucci-Darmanin S,Breuil V等。 成骨细胞中的自噬参与矿化和骨稳态。 自噬。 2014年12月18日; 10(11):1965–77。 6。 Zhao Y,Chen G,Zhang W等。 自噬通过HIF-1α/BNIP3信号通路调节缺氧诱导的破骨细胞生成。 J细胞生理。 2012年2月; 227(2):639–48。 7。 DeSelm CJ,Miller BC,Zou W等。 自噬蛋白调节整骨骨吸收的分泌成分。 DEV单元格。 2011年11月15日; 21(5):966–74。 8。 Sànchez-Riera L,Wilson N,Kamalaraj N等。 骨质疏松和脆弱性骨折。 最佳实践临床风湿性。 9。2008年2月28日; 451(7182):1069–75。2。Mizushima N,Komatsu M.自噬:细胞和组织的翻新。单元格。2011年11月11日; 147(4):728–41。3。Pierrefite-Carle V,Santucci-Darmanin S,Breuil V,Camuzard O,Carle GF。骨骼中的自噬:保持平衡。老化Res Rev.2015年11月; 24(pt b):206-17。 4。 Liu F,Fang F,Yuan H等。 通过抑制成骨细胞末端分化,FIP200缺失对自噬的抑制导致小鼠的骨质减少。 J骨矿工销售J Am Soc Bone Miner Res。 2013年11月; 28(11):2414–30。 5。 Nollet M,Santucci-Darmanin S,Breuil V等。 成骨细胞中的自噬参与矿化和骨稳态。 自噬。 2014年12月18日; 10(11):1965–77。 6。 Zhao Y,Chen G,Zhang W等。 自噬通过HIF-1α/BNIP3信号通路调节缺氧诱导的破骨细胞生成。 J细胞生理。 2012年2月; 227(2):639–48。 7。 DeSelm CJ,Miller BC,Zou W等。 自噬蛋白调节整骨骨吸收的分泌成分。 DEV单元格。 2011年11月15日; 21(5):966–74。 8。 Sànchez-Riera L,Wilson N,Kamalaraj N等。 骨质疏松和脆弱性骨折。 最佳实践临床风湿性。 9。2015年11月; 24(pt b):206-17。4。Liu F,Fang F,Yuan H等。 通过抑制成骨细胞末端分化,FIP200缺失对自噬的抑制导致小鼠的骨质减少。 J骨矿工销售J Am Soc Bone Miner Res。 2013年11月; 28(11):2414–30。 5。 Nollet M,Santucci-Darmanin S,Breuil V等。 成骨细胞中的自噬参与矿化和骨稳态。 自噬。 2014年12月18日; 10(11):1965–77。 6。 Zhao Y,Chen G,Zhang W等。 自噬通过HIF-1α/BNIP3信号通路调节缺氧诱导的破骨细胞生成。 J细胞生理。 2012年2月; 227(2):639–48。 7。 DeSelm CJ,Miller BC,Zou W等。 自噬蛋白调节整骨骨吸收的分泌成分。 DEV单元格。 2011年11月15日; 21(5):966–74。 8。 Sànchez-Riera L,Wilson N,Kamalaraj N等。 骨质疏松和脆弱性骨折。 最佳实践临床风湿性。 9。Liu F,Fang F,Yuan H等。通过抑制成骨细胞末端分化,FIP200缺失对自噬的抑制导致小鼠的骨质减少。J骨矿工销售J Am Soc Bone Miner Res。2013年11月; 28(11):2414–30。5。Nollet M,Santucci-Darmanin S,Breuil V等。 成骨细胞中的自噬参与矿化和骨稳态。 自噬。 2014年12月18日; 10(11):1965–77。 6。 Zhao Y,Chen G,Zhang W等。 自噬通过HIF-1α/BNIP3信号通路调节缺氧诱导的破骨细胞生成。 J细胞生理。 2012年2月; 227(2):639–48。 7。 DeSelm CJ,Miller BC,Zou W等。 自噬蛋白调节整骨骨吸收的分泌成分。 DEV单元格。 2011年11月15日; 21(5):966–74。 8。 Sànchez-Riera L,Wilson N,Kamalaraj N等。 骨质疏松和脆弱性骨折。 最佳实践临床风湿性。 9。Nollet M,Santucci-Darmanin S,Breuil V等。成骨细胞中的自噬参与矿化和骨稳态。自噬。2014年12月18日; 10(11):1965–77。6。Zhao Y,Chen G,Zhang W等。 自噬通过HIF-1α/BNIP3信号通路调节缺氧诱导的破骨细胞生成。 J细胞生理。 2012年2月; 227(2):639–48。 7。 DeSelm CJ,Miller BC,Zou W等。 自噬蛋白调节整骨骨吸收的分泌成分。 DEV单元格。 2011年11月15日; 21(5):966–74。 8。 Sànchez-Riera L,Wilson N,Kamalaraj N等。 骨质疏松和脆弱性骨折。 最佳实践临床风湿性。 9。Zhao Y,Chen G,Zhang W等。自噬通过HIF-1α/BNIP3信号通路调节缺氧诱导的破骨细胞生成。J细胞生理。 2012年2月; 227(2):639–48。 7。 DeSelm CJ,Miller BC,Zou W等。 自噬蛋白调节整骨骨吸收的分泌成分。 DEV单元格。 2011年11月15日; 21(5):966–74。 8。 Sànchez-Riera L,Wilson N,Kamalaraj N等。 骨质疏松和脆弱性骨折。 最佳实践临床风湿性。 9。J细胞生理。2012年2月; 227(2):639–48。7。DeSelm CJ,Miller BC,Zou W等。自噬蛋白调节整骨骨吸收的分泌成分。DEV单元格。2011年11月15日; 21(5):966–74。8。Sànchez-Riera L,Wilson N,Kamalaraj N等。 骨质疏松和脆弱性骨折。 最佳实践临床风湿性。 9。Sànchez-Riera L,Wilson N,Kamalaraj N等。骨质疏松和脆弱性骨折。最佳实践临床风湿性。9。2010年12月; 24(6):793–810。Almeida M,O'Brien CA。 骨骼老化的基本生物学:应力反应途径的作用。 J Gerontol A Biol Sci Med Sci。 2013年10月; 68(10):1197–208。 10。 Manolagas SC,Parfitt AM。 旧的对骨骼意味着什么。 趋势内分泌代替tem。 2010 Jun; 21(6):369–74。 11。 Gavali S,Gupta MK,Daswani B,Wani MR,Sirdeshmukh R,Khatkhatay Mi。 雌激素通过促进自噬来增强人类成骨细胞的存活和功能。 Biochim Biophys acta mol Cell Res。 2019年9月; 1866(9):1498–507。 12。 Cheng L,Zhu Y,Ke D,XieD。雌激素活化的自噬对雌激素的抗αsteocolasogenation具有负面影响。 细胞增殖[Internet]。 2020年3月11日[引用2020年10月12日]; 53(4)。 可从:https://www.ncbi.nlm.nih.gov/pmc/articles/pmc7162800/ 13。 pan F,Liu X-G,Guo Y-F等。 自助途径的调节可能会影响中国的地位变化:老年人的证据。 j hum Genet。 2010年7月; 55(7):441–7。 14。 Zhang L,Guo Y-F,Liu Y-Z等。 基于途径的全基因组关联分析确定了自噬途径对超前半径BMD的重要性。 J骨矿工销售J Am Soc Bone Miner Res。 2010年7月; 25(7):1572–80。 15。 Chen K,Yang Y-H,Jiang S-D,Jiang L-S。 随着衰老的衰老,骨细胞自噬的活性降低可能导致老年人群的骨质流失。 组织化学细胞生物。 16。Almeida M,O'Brien CA。骨骼老化的基本生物学:应力反应途径的作用。J Gerontol A Biol Sci Med Sci。2013年10月; 68(10):1197–208。10。Manolagas SC,Parfitt AM。旧的对骨骼意味着什么。趋势内分泌代替tem。2010 Jun; 21(6):369–74。11。Gavali S,Gupta MK,Daswani B,Wani MR,Sirdeshmukh R,Khatkhatay Mi。雌激素通过促进自噬来增强人类成骨细胞的存活和功能。Biochim Biophys acta mol Cell Res。2019年9月; 1866(9):1498–507。12。Cheng L,Zhu Y,Ke D,XieD。雌激素活化的自噬对雌激素的抗αsteocolasogenation具有负面影响。细胞增殖[Internet]。2020年3月11日[引用2020年10月12日]; 53(4)。可从:https://www.ncbi.nlm.nih.gov/pmc/articles/pmc7162800/ 13。pan F,Liu X-G,Guo Y-F等。自助途径的调节可能会影响中国的地位变化:老年人的证据。j hum Genet。2010年7月; 55(7):441–7。14。Zhang L,Guo Y-F,Liu Y-Z等。 基于途径的全基因组关联分析确定了自噬途径对超前半径BMD的重要性。 J骨矿工销售J Am Soc Bone Miner Res。 2010年7月; 25(7):1572–80。 15。 Chen K,Yang Y-H,Jiang S-D,Jiang L-S。 随着衰老的衰老,骨细胞自噬的活性降低可能导致老年人群的骨质流失。 组织化学细胞生物。 16。Zhang L,Guo Y-F,Liu Y-Z等。基于途径的全基因组关联分析确定了自噬途径对超前半径BMD的重要性。J骨矿工销售J Am Soc Bone Miner Res。2010年7月; 25(7):1572–80。15。Chen K,Yang Y-H,Jiang S-D,Jiang L-S。 随着衰老的衰老,骨细胞自噬的活性降低可能导致老年人群的骨质流失。 组织化学细胞生物。 16。Chen K,Yang Y-H,Jiang S-D,Jiang L-S。随着衰老的衰老,骨细胞自噬的活性降低可能导致老年人群的骨质流失。组织化学细胞生物。16。2014年9月; 142(3):285–95。Camuzard O,Santucci-Darmanin S,Breuil V等。成骨细胞谱系中的性别特异性自噬调制:抵消女性骨质流失的关键功能。oncotarget。2016年10月11日; 7(41):66416–28。17。Yang Y,Zheng X,Li B,Jiang S,Jiang L.卵巢切除大鼠中骨细胞自噬的活性增加,及其与氧化应激状态和骨骼丧失的相关性。Biochem Biophys Res Commun。2014年8月15日; 451(1):86–92。18。Luo D,Ren H,Li T,Lian K,LinD。雷帕霉素通过激活骨细胞自噬来降低老年骨质疏松症的严重程度。骨质骨int j stuph Result coop eur发现了美国的骨质骨骨骨质骨。2016年3月; 27(3):1093–101。19。yuan Y,Fang Y,Zhu L等。 造血自噬的恶化与骨质疏松症有关。 老化细胞。 2020; 19(5):E13114。 20。 Kneissel M,Luong-Nguyen N-H,Baptist M等。 依维莫司通过破骨细胞抑制取消骨质流失,骨吸收和组织蛋白酶K的表达。 骨头。 2004年11月; 35(5):1144–56。 21。 Jia D,O'Brien CA,Stewart SA,Manolagas SC,Weinstein RS。 糖皮质激素直接作用于破骨细胞,以增加其寿命并降低骨密度。 内分泌学。 2006年12月; 147(12):5592–9。 22。 Kim H-J,Zhao H,Kitaura H等。 糖皮质激素通过破骨细胞抑制骨形成。 J Clin Invest。 2006年8月; 116(8):2152–60。 23。 24。yuan Y,Fang Y,Zhu L等。造血自噬的恶化与骨质疏松症有关。老化细胞。2020; 19(5):E13114。20。Kneissel M,Luong-Nguyen N-H,Baptist M等。依维莫司通过破骨细胞抑制取消骨质流失,骨吸收和组织蛋白酶K的表达。骨头。2004年11月; 35(5):1144–56。 21。 Jia D,O'Brien CA,Stewart SA,Manolagas SC,Weinstein RS。 糖皮质激素直接作用于破骨细胞,以增加其寿命并降低骨密度。 内分泌学。 2006年12月; 147(12):5592–9。 22。 Kim H-J,Zhao H,Kitaura H等。 糖皮质激素通过破骨细胞抑制骨形成。 J Clin Invest。 2006年8月; 116(8):2152–60。 23。 24。2004年11月; 35(5):1144–56。21。Jia D,O'Brien CA,Stewart SA,Manolagas SC,Weinstein RS。 糖皮质激素直接作用于破骨细胞,以增加其寿命并降低骨密度。 内分泌学。 2006年12月; 147(12):5592–9。 22。 Kim H-J,Zhao H,Kitaura H等。 糖皮质激素通过破骨细胞抑制骨形成。 J Clin Invest。 2006年8月; 116(8):2152–60。 23。 24。Jia D,O'Brien CA,Stewart SA,Manolagas SC,Weinstein RS。糖皮质激素直接作用于破骨细胞,以增加其寿命并降低骨密度。内分泌学。2006年12月; 147(12):5592–9。22。Kim H-J,Zhao H,Kitaura H等。 糖皮质激素通过破骨细胞抑制骨形成。 J Clin Invest。 2006年8月; 116(8):2152–60。 23。 24。Kim H-J,Zhao H,Kitaura H等。糖皮质激素通过破骨细胞抑制骨形成。J Clin Invest。2006年8月; 116(8):2152–60。 23。 24。2006年8月; 116(8):2152–60。23。24。Lin N-Y,Chen C-W,Kagwiria R等。 自噬的灭活可改善糖皮质激素诱导的卵巢切除术引起的骨质损失。 Ann Rheum Dis。 2016; 75(6):1203–10。 fu L,Wu W,Sun X,ZhangP。糖皮质激素通过PI3K/AKT/MTOR信号通路增强了破骨细胞自噬。 Calcif Tissue int。 2020 Jul; 107(1):60–71。Lin N-Y,Chen C-W,Kagwiria R等。自噬的灭活可改善糖皮质激素诱导的卵巢切除术引起的骨质损失。 Ann Rheum Dis。 2016; 75(6):1203–10。 fu L,Wu W,Sun X,ZhangP。糖皮质激素通过PI3K/AKT/MTOR信号通路增强了破骨细胞自噬。 Calcif Tissue int。 2020 Jul; 107(1):60–71。自噬的灭活可改善糖皮质激素诱导的卵巢切除术引起的骨质损失。Ann Rheum Dis。 2016; 75(6):1203–10。 fu L,Wu W,Sun X,ZhangP。糖皮质激素通过PI3K/AKT/MTOR信号通路增强了破骨细胞自噬。 Calcif Tissue int。 2020 Jul; 107(1):60–71。Ann Rheum Dis。2016; 75(6):1203–10。 fu L,Wu W,Sun X,ZhangP。糖皮质激素通过PI3K/AKT/MTOR信号通路增强了破骨细胞自噬。 Calcif Tissue int。 2020 Jul; 107(1):60–71。2016; 75(6):1203–10。fu L,Wu W,Sun X,ZhangP。糖皮质激素通过PI3K/AKT/MTOR信号通路增强了破骨细胞自噬。Calcif Tissue int。2020 Jul; 107(1):60–71。


凋亡性间充质基质细胞支持骨质核层生成,同时抑制多核巨细胞在体外形成
是由间充质基质细胞(MSC)和磷酸钙(CAP)材料组合诱导的骨再生中的,破骨细胞会作为关键细胞连接炎症和骨形成。 尽管短期植入了植入的MSC,但仍观察到有利的结果,突出了它们的主要旁分泌功能以及细胞死亡在调节其分泌物中的可能影响。 在这项工作中,我们专注于从MSC到整骨细胞的通信。 MSC播种在帽生物材料或经历诱导的凋亡中的 MSC产生了有条件的培养基,该培养基有利于人类CD14+单核细胞的破骨细胞的发展。 相反,MSC的凋亡分泌抑制了IL-4刺激后形成的炎症性多核巨细胞的发展。 使用基于质谱的定量蛋白质组学和主要细胞因子的补充免疫测定法比较了MSC在凋亡应激之前和之后MSC分泌的成分。 CXCR-1和CXCR-2配体,主要是IL-8/CXCL-8,但也建议由生长调节的蛋白CXCL-1,-2或-3作为MSC的主要塑性效应。 这些发现支持以下假设:破骨细胞是骨骼再生的关键参与者,并表明凋亡在MSC的有效性中起着重要作用。,破骨细胞会作为关键细胞连接炎症和骨形成。有利的结果,突出了它们的主要旁分泌功能以及细胞死亡在调节其分泌物中的可能影响。在这项工作中,我们专注于从MSC到整骨细胞的通信。MSC播种在帽生物材料或经历诱导的凋亡中的 MSC产生了有条件的培养基,该培养基有利于人类CD14+单核细胞的破骨细胞的发展。 相反,MSC的凋亡分泌抑制了IL-4刺激后形成的炎症性多核巨细胞的发展。 使用基于质谱的定量蛋白质组学和主要细胞因子的补充免疫测定法比较了MSC在凋亡应激之前和之后MSC分泌的成分。 CXCR-1和CXCR-2配体,主要是IL-8/CXCL-8,但也建议由生长调节的蛋白CXCL-1,-2或-3作为MSC的主要塑性效应。 这些发现支持以下假设:破骨细胞是骨骼再生的关键参与者,并表明凋亡在MSC的有效性中起着重要作用。MSC产生了有条件的培养基,该培养基有利于人类CD14+单核细胞的破骨细胞的发展。相反,MSC的凋亡分泌抑制了IL-4刺激后形成的炎症性多核巨细胞的发展。使用基于质谱的定量蛋白质组学和主要细胞因子的补充免疫测定法比较了MSC在凋亡应激之前和之后MSC分泌的成分。CXCR-1和CXCR-2配体,主要是IL-8/CXCL-8,但也建议由生长调节的蛋白CXCL-1,-2或-3作为MSC的主要塑性效应。这些发现支持以下假设:破骨细胞是骨骼再生的关键参与者,并表明凋亡在MSC的有效性中起着重要作用。

肠道梭状芽胞杆菌衍生的吲哚酸通过激活妊娠X受体
骨稳态通过破骨细胞介导的骨吸收和成骨细胞介导的骨形成保持。绝经后妇女雌激素水平的急剧下降会导致破骨细胞过度活化,骨稳态受损和随后的骨质流失。肠道微生物组的变化会影响骨矿物质密度。但是,肠道微生物组在雌激素缺乏引起的骨质流失及其潜在机制中的作用仍然未知。在这项研究中,我们发现孢子菌的丰度(C. spor。) 及其衍生的代谢产物,吲哚丙酸(IPA)在卵巢切除(OVX)小鼠中降低。 体外测定法表明IPA抑制了破骨细胞的分化和功能。 在分子水平上,IPA抑制了核因子Kappa-配体(RANKL)诱导的妊娠X受体(PXR)泛素化和降解的受体激活剂,从而导致PXR与P65的持续结合增加。 在体内每日IPA给药或重复的C. spor。 定殖侵害了OVX诱导的骨质流失。 保护活细菌免受严峻的胃环境,并延迟口服孢子孢子的排空。 从肠道,一个C. spor。 - 封装的丝纤维蛋白(SF)水凝胶系统,在OVX小鼠中获得了与重复的细菌移植或每日给药相当的OVX小鼠的骨骼保护。 总体而言,我们发现肠道孢子 - 衍生的IPA通过调节PXR/p65复合物来参与雌激素缺乏诱导的破骨细胞过度活化。在这项研究中,我们发现孢子菌的丰度(C. spor。)及其衍生的代谢产物,吲哚丙酸(IPA)在卵巢切除(OVX)小鼠中降低。体外测定法表明IPA抑制了破骨细胞的分化和功能。在分子水平上,IPA抑制了核因子Kappa-配体(RANKL)诱导的妊娠X受体(PXR)泛素化和降解的受体激活剂,从而导致PXR与P65的持续结合增加。在体内每日IPA给药或重复的C. spor。 定殖侵害了OVX诱导的骨质流失。 保护活细菌免受严峻的胃环境,并延迟口服孢子孢子的排空。 从肠道,一个C. spor。 - 封装的丝纤维蛋白(SF)水凝胶系统,在OVX小鼠中获得了与重复的细菌移植或每日给药相当的OVX小鼠的骨骼保护。 总体而言,我们发现肠道孢子 - 衍生的IPA通过调节PXR/p65复合物来参与雌激素缺乏诱导的破骨细胞过度活化。在体内每日IPA给药或重复的C. spor。定殖侵害了OVX诱导的骨质流失。保护活细菌免受严峻的胃环境,并延迟口服孢子孢子的排空。从肠道,一个C. spor。- 封装的丝纤维蛋白(SF)水凝胶系统,在OVX小鼠中获得了与重复的细菌移植或每日给药相当的OVX小鼠的骨骼保护。总体而言,我们发现肠道孢子 - 衍生的IPA通过调节PXR/p65复合物来参与雌激素缺乏诱导的破骨细胞过度活化。C.孢子。包含的SF水凝胶系统是一种有前途的工具,可打击绝经后骨质疏松症,而无需重复的细菌移植。

骨病与治疗
骨质疏松症(OP)是老年常见的退行性疾病,以骨量减少、骨组织结构破坏为特征。骨骼在人体中具有支撑机体、调节代谢、造血等重要作用,在整个机体的生命活动中起着至关重要的作用[1]。因此,骨代谢失衡可能导致OP,进而增加骨折的风险。根据国际骨质疏松基金会的数据,全球每年有超过890万人因OP而骨折,因此对OP疾病的研究至关重要。目前,研究人员正在探索OP的发病机制,寻找更有效的预防和治疗靶点。OP的治疗前景广阔,寻找增加骨形成和维持骨强度的方法,探索治疗靶点在骨发育中的作用及其调控机制,有望成为改善OP疾病的潜在新药靶点。OP是影响老年人生活质量的主要因素之一。临床上,抗骨质疏松药物通常包括促进骨量恢复的骨吸收抑制剂,包括雌激素、降钙素和双膦酸盐 [2]。治疗骨质疏松症的新型药物之一是地诺单抗,这是一种抑制 RANKL 并阻止破骨细胞发育的人源化单克隆抗体。OPG 是一种天然存在的 RANKL 假受体,是一种用于预防和治疗骨质疏松症的基因药物;它与 RANKL 结合以抑制破骨细胞形成,减缓骨量流失并增加骨矿物质密度。OPG 可增加骨矿物质密度并防止小鼠在失重状态下骨骼分解。OPG 已被证明可在小鼠静止不动时防止骨溶解并保持骨骼形成。半胱氨酸蛋白酶、组织蛋白酶 K (Ctsk) 抑制剂(如 Odanacatib (MK0822) 等)等具有骨骼保护作用。成骨细胞和破骨细胞都负责维持动态骨稳态和骨重建[3]。

前列腺癌和骨头:临床表现和分子机制
前列腺癌(PCA)在发达国家中越来越普遍。局部PCA存在有效的治疗选择,但是转移性PCA的治疗选择较少,并且患者的生存率较短。PCA和骨骼健康紧密地交织在一起,因为PCA通常转移到骨架上。由于雄激素受体信号传导驱动PCA生长,因此后遗症降低骨骼强度构成了晚期PCA治疗的基础。骨骼重塑的体内平衡过程 - 由骨建造成骨细胞,骨质骨细胞和调节性骨细胞的一致作用产生。驱动骨骼发育和稳态的机制,例如区域缺氧或基质填充的生长因子,可以被骨转移性PCA征服。以这种方式,维持骨骼的生物学被整合到PCA在骨中生长和存活的自适应机制。由于骨骼生物学和癌症生物学的纠缠性质,骨骼转移性PCA很难进行研究。在此,我们从起源,表现和临床治疗中调查PCA到PCA转移对骨的骨组成以及结构以及分子介质。我们的目的是快速却有效地减少了跨多个学科的团队科学障碍,该学科的重点是PCA和转移性骨病。我们还介绍了组织工程的概念,作为一种新颖的观点,以建模,捕获和研究复杂的癌症微环境相互作用。
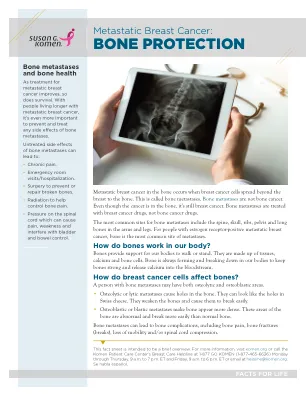
转移性乳腺癌:骨骼保护
骨转移的最常见部位包括脊柱,头骨,肋骨,骨盆和胳膊和腿上的长骨头。对于患有雌激素受体阳性转移性乳腺癌的人来说,骨是最常见的转移部位。骨骼如何在我们的体内起作用?骨骼为我们的身体行走或站立提供了支撑。它们由组织,钙和骨细胞组成。骨头总是在我们体内形成和分解,以保持骨骼坚固并释放到血液中。乳腺癌细胞如何影响骨骼?患有骨转移的人可能具有骨化和成骨细胞区域。

y-box结合蛋白1在小细胞外囊泡中可将间充质干细胞分化为骨细胞 - 急性髓样白血病的刺激性
据报道,急性髓样白血病(AML)细胞释放出的抽象小细胞外囊泡(SEV)会影响骨髓衍生的间充质干细胞的三利分化(BM-MSCS)。但是,AML-Sevs的生物货物对此效果负责,仍然难以捉摸。在这项研究中,使用尺寸排斥色谱和超滤从细胞条件的培养基和血浆中分离出SEV,并根据MISEV2018指南进行表征。我们的结果表明AML-SEVS增加了BM-MSC的增殖。相反,在BM-MSC中下调了对正常造血的关键蛋白。此外,我们揭示了AML-sevs显着降低了BM-MSC对成骨细胞的分化,而不会影响脂肪形成或软骨基因分化。接下来,LC-MS/MS蛋白质组学阐明了包括AML-SEVS处理的AML-SEVS和BM-MSC在包括Y-box结合蛋白1(YBX1)在内的各种蛋白质。在临床上相关,我们发现与健康对照相比,大多数儿科AML衍生的SEV在大多数儿科AML衍生的SEV中都被认为是上调的。有趣的是,在AML细胞中YBX1下调后分离的SEV明显地挽救了BM-MSC的成骨细胞分化。总的来说,我们的数据首次证明了含有AML-SEVS的YBX1是通过降低BM-MSC的骨化分化来破坏骨髓微环境正常功能的关键游戏之一。

buch信息
b细胞 / T细胞(J. E. Konkel,I。L. C. Chapple)胶质细胞和胶质细胞(B. L. Foster,M。Sanz)软骨细胞和纤维软骨细胞(D. S. Nedrelow,M。S. Detamore,M。S. Detamore,M。wong)牙科植物细胞:J。Krivan krivan krivan krivan eker eker eker ek。 (V.-J。Uitto,U。K.Gürsoy)成纤维细胞(G. Pompermaier Garlet,D。S. Thoma)巨噬细胞(J. CW。Wang,W。V. Giannobile)微血管细胞:内皮和周细胞(A. Banfi,S.Kühl)心肌细胞(S. W. Herring,S。Biliaridis)神经细胞(S. B. Oh,P。R. Lee,D。A. Ettlin)Odontlin)Odontlasts(D. D. D. D. D. D. D. Bosshardt,P。R. Schshardl),B。F. Schmidlin(F. Schmidlin)(F. s. fe fe f. s f. s f. s fe fe fe fe fe fe fecoIn of。 Lethaus)骨细胞 / odontoclasts(R。Nishimura,H。Terheyden)骨细胞(R. Gruber,B。Stadlinger)多形核细胞(中性粒细胞)(中性粒细胞)(J. Deschner,S。Jepsen,S。Jepsen)溶性细胞(G. B. B. B. Procter,A。Viss)