XiaoMi-AI文件搜索系统
World File Search System中胞质

tau纤维诱导纳米级膜损伤,并在溶酶体处核定胞质tau
再生或“内在的康复”是指生命系统在受伤或疾病后恢复或康复的能力。也就是说,“治愈”是指恢复,修复或恢复健康,“内在”意味着康复过程在生物体本身内具有其因果关系,而不是通过外部干预依赖因果关系。神经再生有悠久的兴趣史(Stahnisch,2021年),现在对神经退行性疾病和神经系统损伤的再生性相互作用引起了人们的兴趣(Huang等,2021)。当应用于心理健康时,该原则假设具有内在方向性或“目的论”的隐性过程 - 也就是说,它们朝着最终目标迈进,在当前情况下,该目标可能是(非犹太人)康复,恢复或“整体性”(Grof,2012; Vaid&Walker,2022)。隐式,动态的过程将是“ Entelechy”的检验 - 这意味着它们是朝着康复目标迈进的心理和生物学过程。类比通常是在内在的心理和身体康复以及生活系统其他地方的自我调节过程之间进行的(Varela等,1974)。“内在的康复”主题可以在世界各地和整个历史的传统和文化中找到(Campbell,2008年);以及许多整合的健康和健康,精神和宗教习俗,包括瑜伽,心理治疗,呼吸,冥想和祈祷。激活再生机制也与“刺激性”应力(Epel,2020)和“ Jarisch-Herxheimer反应”或“治愈危机”(Bryceson,1976)具有相关性。

环境条件的影响以及胞质谷氨酰胺合成酶对玉米杂化核产生的等位基因变化
胞质谷氨酰胺合成酶(GS1)是主要负责玉米叶中的铵同化和重新合并的酶。通过检查酶在叶细胞中酶的过表达的影响,研究了GS1在玉米核产生中的农艺潜力。使用在该领域生长的植物产生并表征了表现出三倍的叶子GS活性增加三倍的转基因杂种。在不同位置,在叶片和束鞘鞘中的叶片和束鞘鞘中的几种过表达GLN1-3(GLN1-3)的基因(GS1)在不同位置生长了五年。平均而言,与对照组相比,转基因杂种中的核产量增加了3.8%。但是,我们观察到,给定领域试验的环境条件和转基因事件同时依赖于这种增加。尽管从一个环境到另一个环境变化,但在不同位置的两个GS1基因(GLN1-3和GLN1-4)多态性区域和核产量之间也发现了显着关联。我们建议使用基因工程或标记辅助选择的GS1酶是产生高屈服玉米杂种的潜在潜在领导者。但是,对于这些杂种,产量增加将在很大程度上取决于用于种植植物的环境条件。

卟啉单胞菌通过NOD1/KLF5轴调节SCD1依赖性脂质合成
癌症干细胞(CSC)被广泛认为是肿瘤起始和进展的主要介体。近年来,微生物感染与癌症干性之间的关联引起了很大的学术兴趣。卟啉单胞菌(牙龈疟原虫)越来越被认为与口服鳞状细胞癌(OSCC)的发展密切相关。然而,牙龈疟原虫在OSCC细胞的干性中的作用仍然不确定。在此,我们表明牙龈疟原虫与人类OSCC标本中的CSC标记表达呈正相关,促进了OSCC细胞的干性和肿瘤性,并增强了裸鼠的肿瘤形成。从机械上讲,牙龈疟原虫通过上调stearoyl-COA去饱和酶1(SCD1)表达的表达来增加OSCC细胞中的脂质合成,这是一种参与脂质代谢的关键酶,最终导致了茎的获得增强。此外,在体外和体内,OSCC细胞中的SCD1抑制减弱了OSCC细胞的牙龈疟原虫诱导的OSCC细胞的干性,包括CSC标记的表达,球体形成能力,化学耐药性和肿瘤生长。此外,牙龈疟原虫感染的OSCC细胞中SCD1的上调与KLF5的表达相关,并且通过牙龈疟原虫活化的NOD1信号传导调节。在一起,这些发现强调了依赖SCD1依赖性脂质合成在OSCC细胞中的牙龈疟原虫诱导的干性摄取中的重要性,这表明NOD1/KLF5轴可能在调节SCD1表达中起关键作用,并为靶向SCD1作为新的OSCC的靶向SCD1的分子基础。

中国医药大学普通生物学试题
验进行三个世代,且起始dna为100%n 15标记,下列叙述何者最准确地解释世代后的dna分布?(a)第二代中,所有dna第二代中n 14 /n 14 /n 15混合型,第三代出现100%n 14 14型dna(b)dna(b)第一代后的比例占25%,属于轻型DNA的比例占75%
小动物活体光学成像技术在基因和细胞治疗中的应用
基因治疗和递送论文在IVIS上成像1。Agrawal VK,Copeland KM,Barbachano Y,Rahim A,Seth R,White CL,Hingorani M,Nutting CM,Kelly M,Harris P,Pandha H,Melcher AA,Melcher AA,Vile RG,Porter RG,Porter C,Porter C,Harrington KJ。微血管无组织转移用于基因输送:体内评估质粒和腺病毒递送的不同途径。基因治疗。2009年1月; 16(1):78-92。2。ahmed N,Ratnayake M,Savoldo B,Perlaky L,Dotti G,Wels WS,Bhattacharjee MB,Gilbertson RJ,Shine HD,Weiss HL,Rooney CM,Heslop He,Gottschalk S.经过实验性Medulloblastoma的恢复后,HESSCHALK S.经过实验性髓鞘瘤的转移后,具有超含Her2-sperific T细胞的转移。癌症。2007年6月15日; 67(12):5957-5964。3。Ahmed N,Salsman VS,Kew Y,Shaffer D,Powell S,Zhang YJ,Grossman RG,Heslop HE,GottschalkS。Her2特异性T细胞靶向原发性胶质母细胞瘤干细胞并诱导自体实验肿瘤的消退。Clin Cancer Res。 2010年1月15日; 16(2):474-485。 4。 Ahmed N,Salsman vs,Yvon E,Louis Cu,Perlaky L,Wels WS,Dishop MK,Kleinerman EE,Pule M,Pule M,Rooney CM,Heslop HE,GottschalkS。 mol ther。 2009年10月; 17(10):1779-1787。 5。 Akimoto T,Sorg BS,Yan Z.过氧化物酶体增殖物激活的受体 - 伽马共激活剂-1alpha启动子在活小鼠的骨骼肌中的实时成像。 美国生理学杂志,细胞生理学。 2004年9月; 287(3):C790-796。 6。 超声Med Biol。 7。Clin Cancer Res。2010年1月15日; 16(2):474-485。4。Ahmed N,Salsman vs,Yvon E,Louis Cu,Perlaky L,Wels WS,Dishop MK,Kleinerman EE,Pule M,Pule M,Rooney CM,Heslop HE,GottschalkS。 mol ther。 2009年10月; 17(10):1779-1787。 5。 Akimoto T,Sorg BS,Yan Z.过氧化物酶体增殖物激活的受体 - 伽马共激活剂-1alpha启动子在活小鼠的骨骼肌中的实时成像。 美国生理学杂志,细胞生理学。 2004年9月; 287(3):C790-796。 6。 超声Med Biol。 7。Ahmed N,Salsman vs,Yvon E,Louis Cu,Perlaky L,Wels WS,Dishop MK,Kleinerman EE,Pule M,Pule M,Rooney CM,Heslop HE,GottschalkS。mol ther。2009年10月; 17(10):1779-1787。5。Akimoto T,Sorg BS,Yan Z.过氧化物酶体增殖物激活的受体 - 伽马共激活剂-1alpha启动子在活小鼠的骨骼肌中的实时成像。美国生理学杂志,细胞生理学。2004年9月; 287(3):C790-796。6。超声Med Biol。7。Alter J,Sennoga CA,Lopes DM,Eckersley RJ,Wells DJ。微泡稳定性是体内基因转移中介导的超声和微泡效率的主要决定因素。2009年6月; 35(6):976-984。AOI A,Watanabe Y,Mori S,Takahashi M,Vassaux G,Kodama T.使用纳米/微泡和超声波和超声波疱疹疱疹单纯胸腺胸腺胺激酶介导的自杀基因治疗。超声Med Biol。2007年12月18日。8。Arenas F,Hervias I,Uriz M,Joplin R,Prieto J,Medina JF。 ursexyoxycholic和糖皮质激素的组合上调了人肝细胞中AE2替代启动子。 J Clin Invest。 2008年2月; 118(2):695-709。 9。 Asokan A,Johnson JS,Li C,Samulski RJ。 生物发光的病毒粒子壳:定量细胞和活体动物中AAV载体动力学的新工具。 基因治疗。 2008年12月; 15(24):1618-1622。 10。 aung W,Hasegawa S,Koshikawa-Yano M,Obata T,Ikehira H,Furukawa T,Aoki I,Aoki I,SagaT。通过光学和磁共振成像的实验性肿瘤中体内电穿孔介导的转基因表达的可视化。 基因治疗。 2009年7月; 16(7):830-839。 11。 Aung W,Hasegawa S,Koshikawa-Yano M,Tsuji AB,Sogawa C,Sudo H,Sugyo H,Sugyo A,Koizumi M,Furukawa T,SagaT。与Fdg-Pets tumor模型中的可调节性转移基因的表达和评估。 基因治疗。 2010年5月6日。 12。 mol ther。 2009年6月; 17(6):1003-1011。 13。 mol ther。 14。Arenas F,Hervias I,Uriz M,Joplin R,Prieto J,Medina JF。ursexyoxycholic和糖皮质激素的组合上调了人肝细胞中AE2替代启动子。J Clin Invest。2008年2月; 118(2):695-709。9。Asokan A,Johnson JS,Li C,Samulski RJ。生物发光的病毒粒子壳:定量细胞和活体动物中AAV载体动力学的新工具。基因治疗。2008年12月; 15(24):1618-1622。10。aung W,Hasegawa S,Koshikawa-Yano M,Obata T,Ikehira H,Furukawa T,Aoki I,Aoki I,SagaT。通过光学和磁共振成像的实验性肿瘤中体内电穿孔介导的转基因表达的可视化。基因治疗。2009年7月; 16(7):830-839。 11。 Aung W,Hasegawa S,Koshikawa-Yano M,Tsuji AB,Sogawa C,Sudo H,Sugyo H,Sugyo A,Koizumi M,Furukawa T,SagaT。与Fdg-Pets tumor模型中的可调节性转移基因的表达和评估。 基因治疗。 2010年5月6日。 12。 mol ther。 2009年6月; 17(6):1003-1011。 13。 mol ther。 14。2009年7月; 16(7):830-839。11。Aung W,Hasegawa S,Koshikawa-Yano M,Tsuji AB,Sogawa C,Sudo H,Sugyo H,Sugyo A,Koizumi M,Furukawa T,SagaT。与Fdg-Pets tumor模型中的可调节性转移基因的表达和评估。 基因治疗。 2010年5月6日。 12。 mol ther。 2009年6月; 17(6):1003-1011。 13。 mol ther。 14。Aung W,Hasegawa S,Koshikawa-Yano M,Tsuji AB,Sogawa C,Sudo H,Sugyo H,Sugyo A,Koizumi M,Furukawa T,SagaT。与Fdg-Pets tumor模型中的可调节性转移基因的表达和评估。基因治疗。2010年5月6日。12。mol ther。2009年6月; 17(6):1003-1011。13。mol ther。14。Balani P,Boulaire J,Zhao Y,Zeng J,Lin J,WangS。高迁移率组Box2启动子控制的自杀基因表达能够靶向胶质母细胞瘤治疗。Barth AS,Kizana E,Smith RR,Terrovitis J,Dong P,Leppo MK,Zhang Y,Miake J,Olson EN,Schneider JW,Abraham MR,Marban E.带有NA+ CA2+ CA2+ CA2+ CAC2+ CACC2+ CACC2+ CACA2+ CACA2+ CAPIER RECTIER RECTIER CARDICENIC NACSIENIC NICENIC NACCONIC NICEAGIC DEACKICONIC NACELIC NIDEMIAN CARMIDIC NACELIC SACTIIC SACELIC NIDEMIAN IDIAGION的病毒载体。2008年5月; 16(5):957-964。Basile P,Dadali T,Jacobson J,Hasslund S,Ulrich-Vinther M,Soballe K,Nishio Y,Drissi MH,Langstein HN,Mitten DJ,O'Keefe RJ,Schwarz EM,Awad HA。冻干肌腱同种异体移植作为GDF5基因递送的组织工程支架。mol ther。2008年3月; 16(3):466-473。15。Bayer M,Kantor B,Cockrell A,Ma H,Zeithaml B,Li X,McCown T,KafriT。大型U3缺失导致非整合慢病毒载体的体内表达增加。mol ther。2008年12月; 16(12):1968-1976。16。Bell JB,Aronovich EL,Schreifels JM,Beadnell TC,Hackett PB。 的持续时间Bell JB,Aronovich EL,Schreifels JM,Beadnell TC,Hackett PB。

镍基超导体中电荷序的实验研究进展 - 物理学报
图1。ndnio 2中的电荷顺序[24]:(a)从钙钛矿Ndnio 3(灰色)到Infinite-Layer ndnio 2(红色)的还原途径的示意图,具有各种中间状态(蓝色); (b) - (d)样品J的茎结果,可以在面板(d)中区分根尖氧空位,从而导致Q//≈(1/3,0)在傅立叶变换图像(b)中的超晶格峰; (e)在Q //≈(1/3,0)围绕Ni L 3边缘处的弹性RXS测量,实体和虚线分别是具有σ和π偏振入射X射线的数据; (f)在ND M 5边的RXS测量; (g),(h)带有样品C和D的固定波形的RXS信号的能量依赖性,阴影区域表示标称电荷顺序贡献。黑色和红色箭头突出显示了Ni 3D-RE 5D杂交峰和Ni L 3主共振,样品C的中间状态比样品D较大,从而导致超晶格峰更强。

在中质塑料上生物膜形成的程序
微塑料污染已引起公众关注,在某些情况下,甚至被认为是潜在的“行星边界威胁”(Galloway和Lewis,2016; Jahnke等,2017)。在水生环境中,MP在海洋和河流中普遍存在(Horton等,2017; Eriksen等,2017),为水生生物群提供了几种且不断的暴露途径,并有可能通过Ingestion通过Ingestion向人类提供(Boyle等,2020; Senathirajah。; Senathirajah等,20221)。由于其化学性质,MP可以在制造过程中或从促成MPS危害效应的环境中吸附持续的有机污染物(POP),例如多环芳烃(POP)(PAHS)(Gallo等人,2018年)。基于实验室的评估表明,可以进行塑料介导的POP转移到生物体,并且MPS与物质相互作用的机制影响其生物体的生物恢复性以及随之而来的生物积累和生物利用度(Trevisan等人,2019年)。除了这一复杂的过程外,几项研究表明,MP的表面在系统发育和功能上不同的微生物群落中充当人为底物,称为“生物膜”或“表皮界”(Reisser等人,2014年; Zettler等人; Zettler等人,2013年)。表皮微生物群似乎在塑料污染的命运和生态影响中起着关键作用,在过去的几年中,微生物学家正在研究MPS表面上存在的这些社区。该有机层可以充当污染物的储层,影响化学物质的吸附,以吸收对同性恋者生态毒性产生不可预测影响的MPS的生物体的吸附(Rummel等,2017; Flemming等,1995)。由于生物膜吸附特性和降解有机化学物质的能力(Writer等,2011; Wen等,2015),因此在微塑性表面上存在生物膜会影响污染物向生物体的塑性介导的转移。尽管有几项研究表明可能发生污染物的转移(Chua等,2014; Rochman等,2014; Browne等,2013; Gaylor等,2012),但仍不清楚生物膜与塑料相关化学物质的相互作用,从而使其生物利用物与生物体相互作用,并将其与生物体相互作用(and)。因此,该技术报告的目的是提供在微塑性表面上创建生物膜的方法,以便进行中cosmsm实验,可用于评估微塑性相关的生物膜对模型生物体污染物的生物利用度的影响。

挥发性研究中的质谱技术
2不列颠哥伦比亚大学微生物学和免疫学系,加拿大温哥华大学,加拿大温哥华,3个生命科学研究所,不列颠哥伦比亚大学,不列颠哥伦比亚省,不列颠哥伦比亚省,卑诗省,加拿大,加拿大,4 ionicon Analytik geselltik gesellschaft M.B.H.在数据分析中,国家传感器研究中心,都柏林城市大学化学科学学院,都柏林9号,爱尔兰,爱尔兰9号,生物信息学研究生课程,不列颠哥伦比亚大学,不列颠哥伦比亚大学,不列颠哥伦比亚省温哥华大学,加拿大,加拿大8号,8岁不列颠哥伦比亚省温哥华,加拿大10 Ecoscope培训计划,不列颠哥伦比亚大学,不列颠哥伦比亚省温哥华,不列颠哥伦比亚省,加拿大

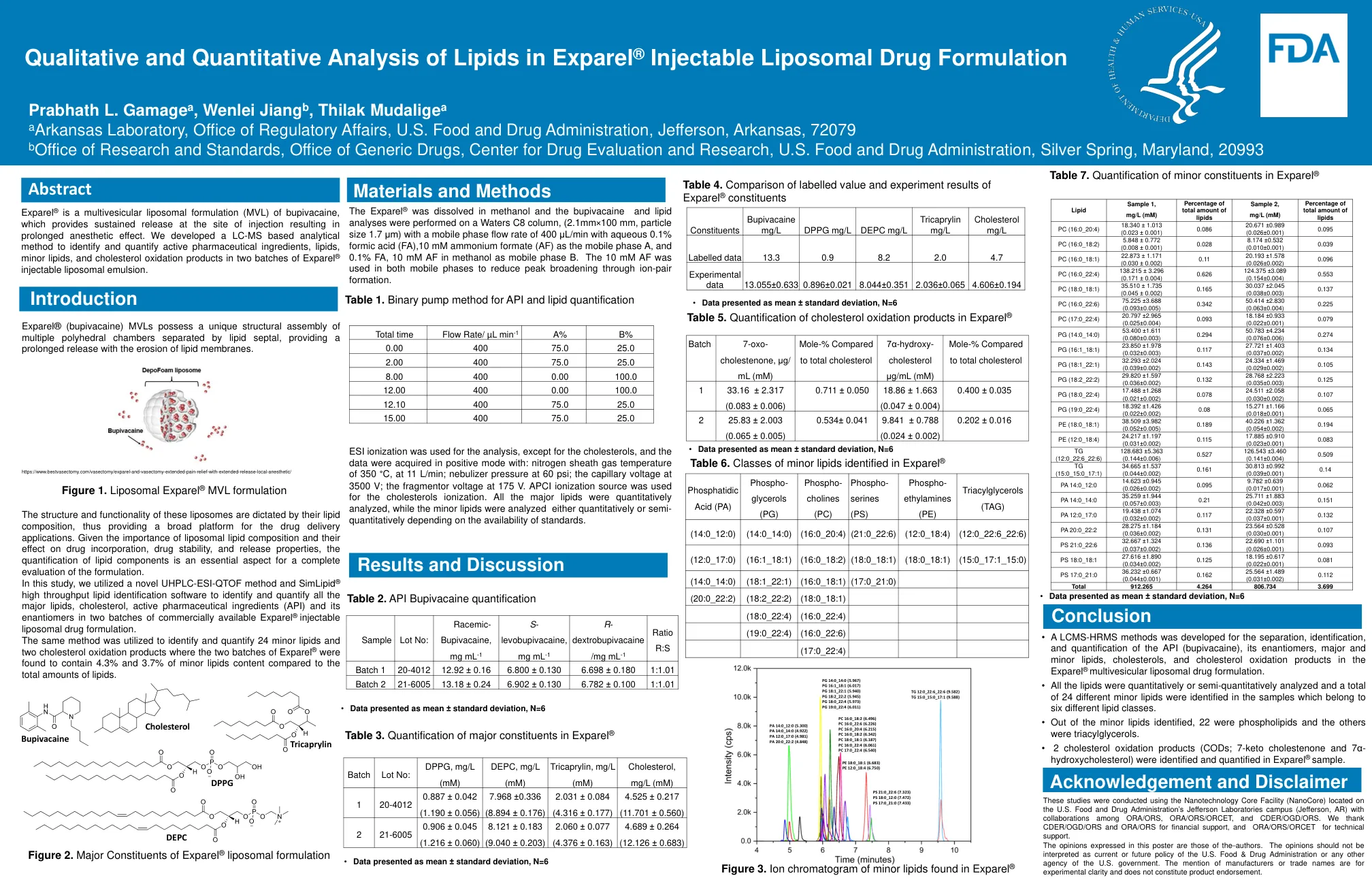
exparel中脂质的定性和定量分析
•开发了一种LCMS-HRMS方法,用于对API(Butivacaine)的分离,鉴定和定量,其对映异构体,主要脂质,胆固醇和胆固醇和胆固醇氧化产物在Exparel®多蛋白脂质体药物制剂中。•对所有脂质进行了定量或半定量分析,并在属于六个不同脂质类别的样品中鉴定了24个不同的小脂质。•从确定的小脂质中,有22个是磷脂,其他是三酰基甘油。•2种胆固醇氧化产物(COD; 7-酮胆固醇和7α-羟基胆固醇)在Exparel®样品中鉴定并量化。