XiaoMi-AI文件搜索系统
World File Search System分泌系统

您需要知道的保护孩子
糖尿病患者被认为是《美国残疾人法案》中的残疾,因为其内分泌系统(一种主要的生活活动)受到了很大的限制。这是说孩子的内分泌系统不起作用的另一种方式,因为它不能正确地制作和/或使用胰岛素。《美国残疾人法》不适用于宗教机构开展的育儿计划。遵守《美国残疾人法案》所需的育儿计划例子是基督教青年会课后计划,城镇经营的夏季营地,国家托儿所特许经营连锁店和睡眠夏季营地。

在Escherichia Coli Nissle 1917中的一种新型益生菌工具包,用于感应和减轻肠道炎症性疾病
摘要:炎症性肠病(IBD)的特征是慢性肠炎,没有治愈和有限的治疗选择,通常具有全身性副作用。在这项研究中,我们开发了一种特定于目标的系统,可以通过设计益生菌大肠杆菌Nissle 1917(ECN)来潜在地处理IBD。我们的模块化系统包括三个组成部分:基于转录因子的传感器(NORR),能够检测炎症生物标志物一氧化氮(NO),1型血素蛋白分泌系统以及由人类抗TNFα纳米型的库组成的治疗货物。尽管敏感性降低,但我们的系统表现出对NO的浓度依赖性反应,成功地分泌了与常用药物adalimumab相当的结合亲和力的功能性纳米型,如酶联免疫吸收测定和体外分析所证实。这个新验证的纳米库库扩展了ECN治疗功能。也可以在ECN中首次表征所采用的分泌系统,可以进一步改编为筛选和净化感兴趣的蛋白质的平台。此外,我们提供了一个数学框架来评估工程益生菌系统中的关键参数,包括相关分子的产生和扩散,细菌定植率和粒子相互作用。这种综合方法扩展了用于基于ECN的疗法的合成生物学工具箱,提供了新颖的零件,电路和炎症热点可调反应的模型。关键字:工程益生菌,IBD,渗透性,E。Coli Nissle 1917(ECN),一氧化氮,TNFα,纳米型■简介

处女座,Mollie和Mostowy,S。和Ho,Brian(2024)使用斑马鱼来识别特定于类型VI分泌系统介导的Interbact Interbact
细菌间竞争会塑造宿主中发现的微生物群落,但是这场比赛与宿主防御之间的相互作用尚不清楚。在这里,我们使用斑马鱼后脑心室(HBV)作为体内平台,以研究具有不同形式的细菌间竞争形式的定义细菌群落的宿主反应。我们发现,来自Vibrio Cholerae和Acinetobacter baylyi的VI型分泌系统(T6SS)的抗菌活性都可以诱发宿主炎症,并使宿主敏感到独立于任何个体效应子的感染。化学抑制炎症可以解决宿主存活中T6SS依赖性差异,但是两种细菌物种之间发生这种情况的机制有所不同。相比之下,尽管志贺氏菌sonnei菌株是一种更有效的细菌杀手,但引起了大结菌素介导的拮抗作用,导致宿主的反应可忽略不计,导致对A的影响没有影响。baylyi或v。霍乱毒力。总的来说,这些结果提供了有关体内不同模式的不同模式如何以不同的方式影响宿主的方式。

依赖T6SS依赖性效应子RE78 ETLI MIM1有益细菌竞争
简单摘要:根瘤菌ETLI MIM1(REMIM1)具有活性在自由生活和共生中的VI型蛋白质分泌系统。T6SS是一种纳米芳烃,将称为效应子的蛋白质分泌为真核和原核靶细胞。REMIM1 T6SS基因簇编码有毒效应子(RE78)以及免疫蛋白(RE79),如在大肠杆菌中表达时所证明的。另外,观察到RE78蛋白的毒性作用在细胞质之外,因为仅当将信号肽添加到其中时才发生对大肠杆菌的毒性作用。RE79在Remim1 Periplasm中发现,并且与T6SS的易位无关。此外,RE78/RE79对还参与细菌竞争和结节占用率。更好地理解该分泌系统的作用对于选择高度竞争性根茎的接种剂可能非常有用。

细菌间 DNA 脱氨酶毒素直接诱变幸存的目标种群
摘要 当细菌细胞接触时,通常会通过毒素传递介导拮抗作用。此类接触对受体细胞产生长期有益影响的可能性尚未得到研究。在这里,我们研究了 DddA 中毒的影响,DddA 是一种胞嘧啶脱氨酶,通过伯克霍尔德菌的 VI 型分泌系统 (T6SS) 传递。尽管 DddA 具有杀灭潜力,但我们观察到几种细菌对 DddA 有抵抗力,反而会积累突变。这些突变可导致获得抗生素耐药性,这表明即使在没有杀灭的情况下,细菌间拮抗作用也会对目标群体产生深远影响。对脱氨酶超家族中其他毒素的研究表明,诱变活性是这些蛋白质的共同特征,包括我们展示的代表性毒素,它以单链 DNA 为目标,并显示出明显不同的结构。我们的研究结果表明,细菌间拮抗相互作用的一个令人惊讶的结果可能是通过直接诱变毒素的作用促进适应。
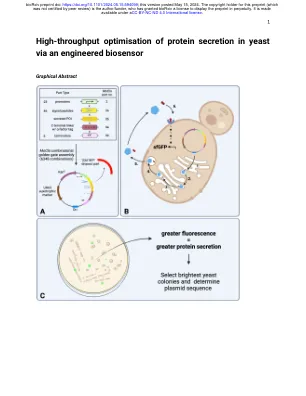
通过工程生物传感器
高价值蛋白质和酶的分泌是合成生物学经济的基础;允许在生产过程中连续发酵和蛋白质纯化而无需细胞裂解。大多数真核蛋白分泌由N末端信号肽编码;但是,信号肽序列变化对给定蛋白的分泌效率的强大影响尚未很好地定义。尽管天然信号肽序列多样性高,但大多数重组蛋白分泌系统仅采用少数表征良好的信号肽。此外,启动子和终止剂的选择可以显着影响分泌效率,但筛选众多遗传构建体以使最佳序列效率低下。在这里,我们调整了酵母G蛋白偶联受体生物传感器,以测量与任何感兴趣蛋白共归因于肽的肽标签的浓度。蛋白质分泌效率可以通过诱导受体激活下游的荧光报告基因来量化。此功能可以使用一锅组合金门克隆组装,对6000多个启动子,信号肽和终结器的6000多个组合进行高通量筛选。我们证明了这种生物传感器可以快速识别分泌和量化分泌水平的最佳组合。

肽聚糖编辑在细菌战中为鲍曼不动杆菌提供免疫力
肽聚糖 (PG) 在大多数细菌中是必需的。因此,它经常成为各种攻击的目标,包括通过 VI 型分泌系统 (T6SS) 的细菌间攻击。本文中,我们报告了革兰氏阴性细菌鲍曼不动杆菌菌株 ATCC 17978 在稳定期产生、分泌并将非规范 d -氨基酸 d -赖氨酸掺入其 PG 中。我们表明,PG 编辑通过提供针对各种细菌竞争对手的肽聚糖靶向 T6SS 效应物的免疫力,提高了鲍曼不动杆菌在细菌战中的竞争力。相反,我们发现 d -Lys 的产生不利于致病机制,至少部分原因是人类酶 d -氨基酸氧化酶 (DAO) 的活性,它会降解 d -Lys 并产生对细菌有毒的 H 2 O 2。系统发育分析表明,鲍曼不动杆菌的最后共同祖先具有产生 d -Lys 的能力。然而,这种特性已独立丧失多次,可能反映了鲍曼不动杆菌作为人类病原体的进化。

遗传和功能多样性有助于解释Xanthomonas属的致病性,弱致病性和共同生活方式
Xanthomonas属主要研究了与植物的致病相互作用。然而,除了宿主和TIS特异性的致病菌株外,该属还包括从广泛宿主分离的非pt造菌株,有时与致病性菌株和其他环境有关,包括雨水。基于它们的丧失能力或有限的能力在隔离宿主上引起症状的能力有限,非对Xanthomonads可以进一步将其描述为共生和弱致病性。这项研究旨在根据其基于其同时发生和系统发育关系的致病性对应物,了解非对照性黄金元中的多样性和演变,并以生态策略的形式构成了生命历史框架的基因组性状。我们测序了跨越系统发育的83个菌株的基因组,并鉴定出8种新型物种,表明未开发的多样性。尽管某些非致病性物种最近损失了III型分泌系统,特别是HRP2群集,但我们观察到HRP2群集与各种物种的生活方式显然缺乏关联。,我们对337个Xanthomonas菌株的大量数据集进行了关联分析,以解释黄thomonads如何成为与植物的社会化,从共生到弱病原体到病原体的植物。存在明显的转录调节剂,不同的营养利用和同化基因,转录调节剂和化学出租车基因可能解释了Xanthomonads的生活方式特异性适应性。

来自巴西坚果树的Pantoea Stewartii RON18713的完整基因组序列揭示了参与植物生长促进的基因
摘要:亚马逊雨林是物种数量和众多的近卫生关系中的超多样性生态系统。为了表征占主导地位和经济重要的亚马逊物种,巴西坚果树(Bertholletia Excelsa Bonpl。),在基因组水平上,从单个个体的叶子中引发了高覆盖的长阅读测序数据。基因组组装揭示了一个意外的发现:两个可以分配给染色体的圆形重叠群和pantoea stewartii菌株的质粒。比较基因组学表明,该菌株属于独立元素亚种,并与从新热带棕榈bactris gasipaes kunth的患病叶片中分离出的其他菌株高度同步。对致病性相关基因的研究揭示了质粒中没有整个III型分泌系统基因簇,质粒否则与已知在Dracaena Sanderiana Mast中引起疾病的分离物的质粒高度相似。相反,检测到与植物生长有关的几种基因,包括参与吲哚-3-乙酸(IAA)产生的基因,磷酸盐溶解和辅助载体的生物合成。总而言之,我们报告了未经培养的Stewartii亚种的基因组。与巴西坚果树相关的植物菌株,并可能是植物生长的细菌。

加强学习中的概括
幽门螺杆菌的惊人特征之一是临床分离株之间广泛的遗传多样性。这种多样性归因于突变率升高,DNA修复受损,DNA转移和频繁重组事件。质粒也已在幽门螺杆菌中鉴定出来,但尚不清楚连接是否可以导致临床分离株之间的DNA转移。检查幽门螺杆菌是否具有共轭质粒转移的固有能力,将穿梭载体引入了幽门螺杆菌中,其中含有含有共轭Incp质粒质粒RP4的原始序列,但没有动员(MOB)基因。表明,这些载体可以稳定复制并在幽门螺杆菌菌株中动员。还证明,幽门螺杆菌染色体上携带的trag和弛豫酶(RLX)同源物对于质粒转移至关重要。引物扩展研究和诱变进一步证实了幽门螺杆菌中的弛豫酶同源物RLX1编码能够在RP4 ORIT上作用的功能酶。此外,这项研究的发现表明,TRAG和RLX1独立于先前描述的IV型分泌系统,包括由CAG致病性岛和梳子转化设备编码的,在介导H. Pylori菌株之间的结合质粒DNA转移中。