XiaoMi-AI文件搜索系统
World File Search System密码子

尿苷RNA在烟草中编辑(Nicotiana
四个组织,而烟草根,茎,叶和花中组织特异性编辑位点的数量相应为5、21、17和35(表S4中列出了详细信息)。同时,有5个组织特异性的编辑基因。两个基因ORF306和ORF151分别在茎和叶中进行了专门编辑,而三个基因细胞色素C成熟基因(CCMB,CCMFC)和30S核糖体基因(RPS14)在花中专门编辑。只有ORF151基因具有两个RNA编辑位点,而其余的四个基因仅具有一个位点。 ,这些基因中的所有编辑位点都位于密码子的第二个位置。 通过比较各种组织中RNA编辑位点的分布(补充图S2),我们观察到NADH脱氢酶亚基的编辑事件在根中显然降低了,这与先前的研究结果一致(Chateigner-只有ORF151基因具有两个RNA编辑位点,而其余的四个基因仅具有一个位点。,这些基因中的所有编辑位点都位于密码子的第二个位置。通过比较各种组织中RNA编辑位点的分布(补充图S2),我们观察到NADH脱氢酶亚基的编辑事件在根中显然降低了,这与先前的研究结果一致(Chateigner-

神经tRNA修饰疾病的分子机制 - 生物化学
1)Suzuki,T。(2021)tRNA修改的扩展世界及其疾病相关性。nat。修订版mol。细胞生物。 ,22,375 - 392。 2)Chujo,T。&Tomizawa,K。(2021)人类转移RNA模量:由转移RNA修改中的畸变引起的疾病。 febs J.,288,7096 - 7122。 3)Asano,K.,Suzuki,T.,Saito,A.,Wei,F.-Y.,Ikeuchi,Y.,Numata,T.,Tanaka,R.,tanaka,R.,Yamane,Y. (2018)与牛磺酸降低和人类疾病相关的tRNA修饰的代谢和化学调节。 核酸res。 ,46,1565 - 1583。 4) (2011)CDKAL1对TRNA(LYS)修饰的词置换会导致小鼠2型糖尿病的发展。 J. Clin。 投资。 ,121,3598 - 3608。 5) (2021)FTSJ1的损失渗透了大脑中特定的翻译效率,并且与X连锁的智力障碍有关。 SCI。 adv。 ,7,EABF3072。 6)Tresky,R.,Miyamoto,Y.,Nagayoshi,Y.,Yabuki,Y.,Araki,K.,Takahashi,Y.,Komohara,Y. (2024)TRMT10A功能障碍Perturbs密码子蛋氨酸和谷氨酰胺的平移,并损害小鼠的脑功能。 nucl。 酸res。细胞生物。,22,375 - 392。2)Chujo,T。&Tomizawa,K。(2021)人类转移RNA模量:由转移RNA修改中的畸变引起的疾病。febs J.,288,7096 - 7122。3)Asano,K.,Suzuki,T.,Saito,A.,Wei,F.-Y.,Ikeuchi,Y.,Numata,T.,Tanaka,R.,tanaka,R.,Yamane,Y.(2018)与牛磺酸降低和人类疾病相关的tRNA修饰的代谢和化学调节。核酸res。,46,1565 - 1583。4)(2011)CDKAL1对TRNA(LYS)修饰的词置换会导致小鼠2型糖尿病的发展。J. Clin。 投资。 ,121,3598 - 3608。 5) (2021)FTSJ1的损失渗透了大脑中特定的翻译效率,并且与X连锁的智力障碍有关。 SCI。 adv。 ,7,EABF3072。 6)Tresky,R.,Miyamoto,Y.,Nagayoshi,Y.,Yabuki,Y.,Araki,K.,Takahashi,Y.,Komohara,Y. (2024)TRMT10A功能障碍Perturbs密码子蛋氨酸和谷氨酰胺的平移,并损害小鼠的脑功能。 nucl。 酸res。J. Clin。投资。,121,3598 - 3608。5)(2021)FTSJ1的损失渗透了大脑中特定的翻译效率,并且与X连锁的智力障碍有关。SCI。 adv。 ,7,EABF3072。 6)Tresky,R.,Miyamoto,Y.,Nagayoshi,Y.,Yabuki,Y.,Araki,K.,Takahashi,Y.,Komohara,Y. (2024)TRMT10A功能障碍Perturbs密码子蛋氨酸和谷氨酰胺的平移,并损害小鼠的脑功能。 nucl。 酸res。SCI。adv。,7,EABF3072。6)Tresky,R.,Miyamoto,Y.,Nagayoshi,Y.,Yabuki,Y.,Araki,K.,Takahashi,Y.,Komohara,Y.(2024)TRMT10A功能障碍Perturbs密码子蛋氨酸和谷氨酰胺的平移,并损害小鼠的脑功能。nucl。酸res。,52,9230 - 9246。7)Blanco,S.,Dietmann,S.,Flores,J.-V.,Hussain,S.,Kutter,C.,Humphreys,P.,Lukk,M.,Lombard,P.,Treps,L.,Popis,M。等。(2014)TRNA的异常甲基化将细胞应激与神经发育疾病联系起来。Embo J.,33,2020 - 2039。

1740750805.pdf
当Tra2β蛋白水平太高时,UCE会触发基因RNA中的额外外显子,引入了停止蛋白质合成的终止密码子,防止过度生产。突变破坏UCE的蛋白质限制功能会导致不育症,从而阻止遗传。因此,自然选择已在数百万年内保留了整个物种的UCE。超保存的元素:UCE是至少200个碱基对的脱氧核糖核酸(DNA)序列,它们在多种物种中一直保持不变,持续了8000万年或更长时间。

自由生活啮齿动物中的细胞因子基因多态性和寄生虫敏感性:非编码变体的重要性
遗传变异与感染易感性之间的关联长期以来一直在自由宿主中进行研究,以推断出塑造免疫基因遗传多态性的当代进化力。尽管对蛋白质与病原体衍生的配体相互作用,例如MHC(主要的组织相互作用复合物)或TLR(TLR样受体),但对免疫系统的传递臂知之甚少。细胞因子是触发和调节免疫反应的信号分子,是先天性和适应性免疫之间的关键联系。In the present study we investigated how genetic variation in cytokines in bank voles Myodes glareolus affects their susceptibility to infection by parasites (nematodes: Aspiculuris tianjensis , Heligmosomum mixtum , Heligmo- somoides glareoli ) and microparasites ( Cryptosporidium sp , Babesia microti , Bartonella sp . )。我们专注于三种细胞因子:肿瘤坏死因子(TNF),淋巴毒素α(LTα)和干扰素β(IFNβ1)。总体而言,我们确定了与线虫易感性相关的四个单核苷酸聚合物(SNP):两个位于ltα中的两个单核苷酸(SNP),其中两个位于IFNβ1中。其中一个变体是代名词,另一个位于内含子中。与寄生虫负载相关的每个SNP都位于选择的密码子中或旁边,三个密码子显示了阳性选择的签名,以及一个净化选择之一。我们的结果表明,细胞因子容易受到寄生虫驱动的选择,而非编码变体虽然在宿主寄生虫共同进化的遗传背景中通常被忽略,但可能在野生系统中感染的易感性中起作用。

引用本文:Aboulalaa K、Laraqui A、Tagajdid R、Ennibi K、Ennaji MM。TP53 基因的种系突变可能是前列腺癌发生的关键因素
目的:本研究旨在调查在已知患有前列腺癌的男性血液中是否可以检测到 TP53 基因外显子 5 的种系变异,并评估影响该基因的基因组变异与患者临床病理特征之间的潜在关联。方法:对 48 例确诊为前列腺癌的男性血液样本进行 TP53 种系突变分析,并通过 Sanger 测序进行确认。根据患者的病理标准分析高频突变的频率和分布,并进行计算研究以评估新突变的影响。结果:Sanger 测序显示,79% 的研究人群携带 TP53 基因突变。总之,该基因共鉴定出 137 种突变,其中 115 种是新突变。移码突变最为常见;15 例(31%)记录了 c.392delA 突变;突变 c.383delC 和 c.432delG 的频率分别为 12.5% 和 10%。最常见的错义突变是变体 c.502C>A (p.His168Asn),发生在 11 名患者 (23%) 中。在一名患者中发现了一个无义突变,导致 126 位 (酪氨酸) 出现终止密码子。受这些改变影响的所有密码子都是蛋白质 TP53 的 DNA 结合域的一部分。结论:在前列腺癌患者中观察到的种系突变频率和 TP53 基因中记录的新突变可能支持该基因的基因组改变与前列腺癌发生之间存在潜在关联,从而构成一种工具,类似于 DNA 修复途径中的其他基因,例如 BRCA1 和 BRCA2。这可能有助于前列腺癌诊断和治疗策略的进步。
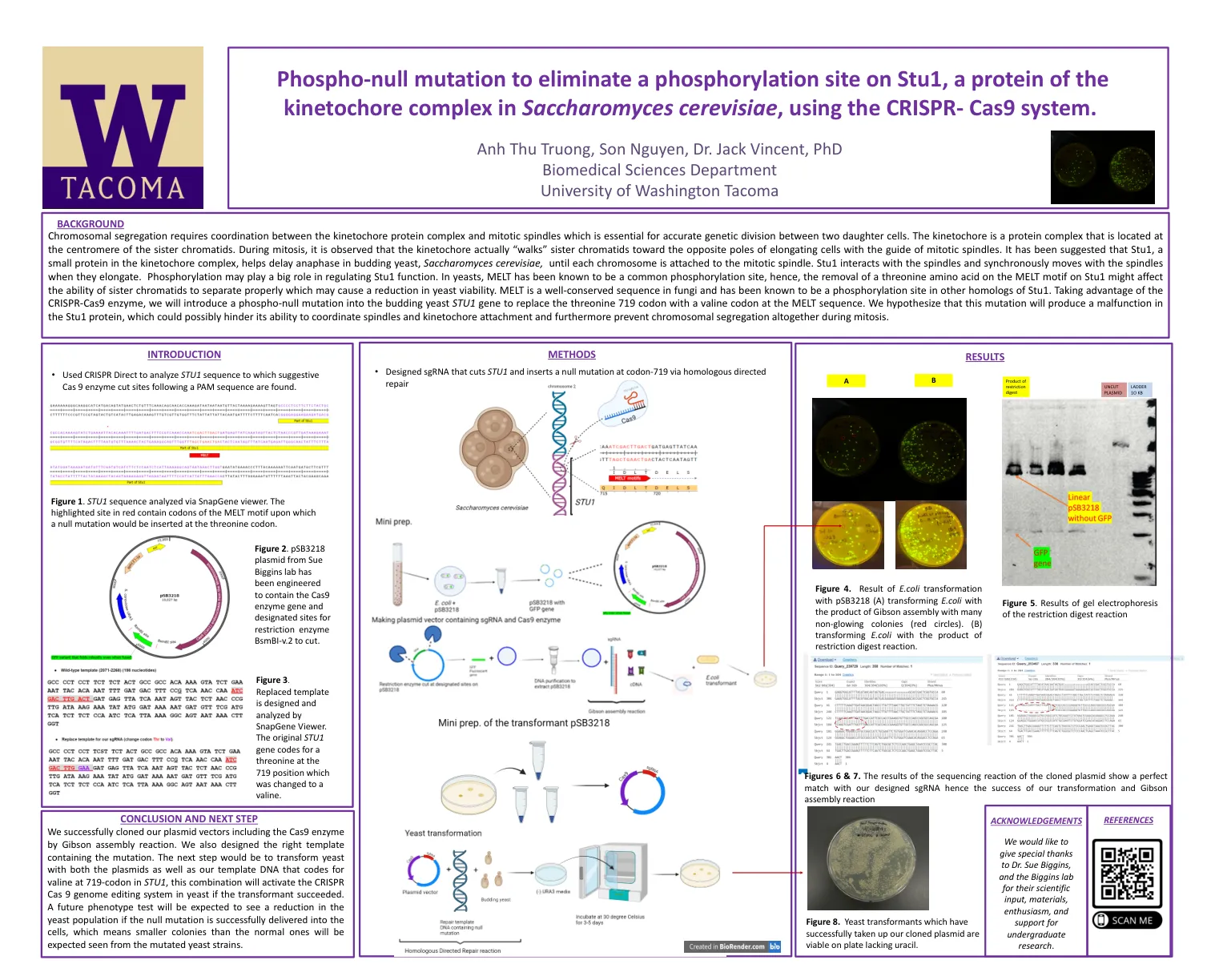
利用 CRISPR 技术进行磷酸化无效突变,消除酿酒酵母动粒复合体蛋白 Stu1 上的磷酸化位点
染色体分离需要动粒蛋白复合物和有丝分裂纺锤体的协调,这对于两个子细胞之间的准确遗传分裂至关重要。动粒是一种位于姊妹染色单体着丝粒的蛋白复合物。在有丝分裂过程中,可以观察到动粒实际上是在有丝分裂纺锤体的引导下将姊妹染色单体“引导”到伸长细胞的相反极点。有人提出,动粒复合物中的小蛋白 Stu1 有助于延迟芽殖酵母酿酒酵母的后期,直到每条染色体都附着在有丝分裂纺锤体上。Stu1 与纺锤体相互作用,并在纺锤体伸长时与其同步移动。磷酸化可能在调节 Stu1 功能方面发挥重要作用。在酵母中,MELT 是一种常见的磷酸化位点,因此,去除 Stu1 上 MELT 基序上的苏氨酸氨基酸可能会影响姐妹染色单体正确分离的能力,从而导致酵母活力下降。MELT 是真菌中保存良好的序列,并且已知是 Stu1 其他同源物中的磷酸化位点。利用 CRISPR-Cas9 酶,我们将在芽殖酵母 STU1 基因中引入磷酸化无效突变,以将 MELT 序列中的苏氨酸 719 密码子替换为缬氨酸密码子。我们假设这种突变会导致 Stu1 蛋白发生故障,这可能会阻碍其协调纺锤体和着丝粒附着的能力,并进一步阻止有丝分裂期间染色体分离。

国际寄生虫学杂志
Haemonchus contortus是小型反刍动物中最致病的线虫,而驱虫抗性(AR)阻碍了其有效的控制。需要早期检测AR状态才能减少AR的选择,并且无法使用表型测试来实现。对于苯二唑唑(BZ),在同种型1β-微型蛋白基因中以单核苷酸多态性(SNP)为特征的AR相关等位基因的检测允许Stron Gyles的早期AR检测。在抗BZ的种群中已经描述了F200Y,F167Y,E198A和E198L多态性,区域之间的频率有明显变化。一种新型的数字PCR(DPCR)可以检测H. contortus中所有上述多态性。测定进行了验证。然后,分析了26个奥地利人和10个意大利绵羊农场的幼虫,并在农场一级合并。对于所有测定,证明了15份/μL电阻等位基因的检测极限和高度准确性,从而可以在大多数样品中检测1%的等位基因频率。在奥地利的样本中,在所有农场都检测到了F200y等位基因的频率升高。第一次在奥地利的H. tortus中发现了密码子167和密码子198中的多态性。在意大利样品中,电阻等位基因的频率仍然相对较低,但F200Y抗性等位基因可追溯。总而言之,我们首次开发了DPCR分析,该测定目标是针对H. contortus中与BZ抗性相关的所有相关性SNP。对AR开发的未来研究可能会受益于基于SNP的监视,其中包括所有相关性SNP的开发测定法。改进的监视将包括其他重要的,尽管病原体较少的线虫属。

单次静脉注射合成外泌体递送的 CRISPR 基因后,成功将阿尔茨海默病模型小鼠脑中的载脂蛋白 E4 编辑为 E3
图 1 . ApoE4 gRNA 变体、PAM 位点和 E4 ARG 到 E3 CYS 的碱基编辑。(A)显示的是 gRNA #1 和 #2,它们以 APOE4 序列“C”为目标,并分别在位置 #8 和 #5 处将其与胞嘧啶脱氨酶胞嘧啶编辑窗口对齐(框出)。密码子 112 中的“C”到“T”碱基编辑导致 ARG 到 CYS 替换,从而产生 ApoE3。(B)显示了 CBE(Cas9n、evoAPOBEC1 和 UGI)、E4 特异性 gRNA 和 sgRNA 以及 ApoE4 基因复合物。目标胞嘧啶“C”显示在胞嘧啶脱氨酶 evoAPOBEC1 酶附近。选择了两个候选 CBE,pBT375 和 pYE1BE4max,34 进行合成和测试,

寡转移和寡进展性疾病的治疗:最新进展和前瞻性临床研究 - 结直肠癌和
所示细胞系用浓度逐渐增加的西妥昔单抗处理 4 天,并通过测量 ATP 含量来评估细胞活力。条形图代表西妥昔单抗对每种细胞系影响的任意指数,如方法中所述。对西妥昔单抗敏感的细胞系显示为负指数。红色条形图代表 KRAS 改变的细胞系;黄色条形图表示 NRAS 突变细胞;蓝色条形图表示影响 BRAF 密码子 V600 的基因改变;黑色条形图表示 RAS/BRAF 野生型细胞。NCIH630 细胞是 KRAS 扩增的46

微藻基因工程的最新进展
图 1. 开发微藻作为商业产品生物制造平台的遗传工具。生物信息学算法用于分析藻类基因组序列,从而产生密码子优化和基序发现技术,这些技术允许设计用于藻类菌株遗传转化的强表达载体。启动子和转录因子等调控元件允许重组基因表达和代谢途径操纵以获得感兴趣的产品。随机诱变和基因组改组可以进一步推动藻类生产菌株向所需的表型发展。这些工具正被用于探索从微藻中工业化生产食品、燃料、材料和药物。