XiaoMi-AI文件搜索系统
World File Search System屏状核

在人造细胞内创建模拟细胞核的分隔结构
这项研究得到了日本科学技术振兴机构 (JST) 战略基础研究促进计划 CREST“用于长 DNA 合成和自主人工细胞创建的人工细胞反应器系统”研究领域 (编号 JPMJCR19S4)、GteX“大规模并行蛋白质打印机系统的开发”研究领域 (编号 JPMJGX23B1)、ASPIRE“日英合作开发人工光合细胞系统”(编号 JPMJAP24B5) 和科学研究补助金“Kikagaku S”(编号 JP19H05624) 的支持。 术语表(注1) 真核生物:具有细胞核并被核膜包围,且含有线粒体等细胞器的生物的统称。它们包括动物、植物和真菌,具有比原核生物更复杂的细胞结构。 (注2)内在无序蛋白质是在生理条件下不能形成三维结构的蛋白质,与酶等折叠成特定的三维结构才能发挥功能的蛋白质不同。分子间多样化的相互作用网络推动液-液相分离,形成称为凝聚层的液滴。 (注3)液-液相分离:均质液体混合物自发分离成两个具有不同成分的液相的现象。单一聚合物(如天然存在的变性蛋白质)可发生相分离,形成致密相和稀相,或者两种不同组成的致密相(如葡聚糖和聚乙二醇)。 (注4)肽标签:一种用于连接特定蛋白质的短氨基酸序列。通过将DNA序列遗传整合到蛋白质中,可以很容易地将其添加到蛋白质中。本研究中使用的肽标签具有拉链式结构,使得它们能够相互互锁并进行特定结合。另一方面,由于它几乎不与其他分子或蛋白质结合,因此可以利用这一特性选择性地将特定蛋白质结合在一起。在该系统中,一个肽标签附着在IDP上,另一个肽标签附着在要掺入IDP相的蛋白质上。 (注5)分子信标:用于检测特定DNA或RNA序列的核酸探针,具有包含荧光染料和猝灭剂的环状结构。在没有目标序列的情况下,荧光就不会出现,但一旦与序列结合,分子的形状就会发生变化,发出荧光并变得可检测。这可以实时确认样本中特定基因或 RNA 的存在。


基于增材制造的新型制造
1)Wohlers, T.:Wohlers Report 2005, p.157, Wohlers Associate Inc., CO, USA(2005 年) 2)https://www.aligntech.com/solutions(访问日期 2020/02/24) 3)Imagawa, Edagawa 等:Phys. Rev. B, 82(11),115116(2010 年) 4)Niino, Hamajima 等:Biofab, 3(3),034104(2011 年)

人类上核核中脑活动的昼夜变化
研究文章|人类脑活动的系统/电路在人类上部核中https://doi.org/10.1523/jneurosci.1730-23.2024收到:2023年9月13日被修订:2023年11月29日接受:2024年1月9日,2024年1月9日,2024年1月29日,授权
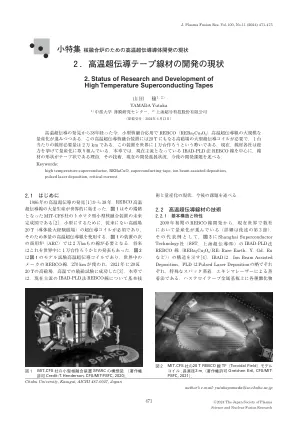
2。高温超导胶带电线的发展现状
[15] Watanabe Tomonori等人:低温工程39,553(2004)。[16] Iimi Akira等人:低温工程42,42(2007)。[17] A.P.Malozemoff和Y. Yamada:超导100年,第11章“第二代HTS Wire”,P689(CRC出版社,2011年)。和Izumi Teruro,Yanagi Nagato:血浆和核融合杂志93,222(2017)。大量的制造方法,包括兔子底物,mod(化学溶液方法)和真空蒸发方法。 [18] http:// www。istec。或。JP/Tape-Wire/Labo-Tape-Wire。html,使用PLD方法和MOD方法(化学溶液方法)的金属棒的高性质。[19] T. Haugan等。,自然430,867(2004)。[20] Y. Yamada等。,应用。物理。Lett。 87,132502(2005)。 [21] H. Tobita等。 ,超级条件。 SCI。 技术。 25,062002(2012)。 [22] Matsumoto Kaname:应用物理77,19(2008)。 [23] Yamada Shigeru:应用物理93,206(2024)。 [24] Y. Yamada,第36届国际超导性国际研讨会(ISS2023),Takina,新西兰惠灵顿,11月28日至30日,2023年。 [25] Miyata Noboru:材料37,361(1988)。 [26] https://www.t.u-tokyo.ac.jp/press/pr2023-06-28-001 [27] A. Stangl等。 ,科学。 Rep。11,8176(2021)。 [28] R. Hiwatari等。 ,血浆融合res。 14,1305047(2019)。 [29]在美国休斯顿大学申请2023年国际申请指挥的布兰登·索博姆(Brandon Sorbom)(2023年)。 [30] D. uglietti,超越。 SCI。 技术。 32,053001(2019)。Lett。87,132502(2005)。[21] H. Tobita等。,超级条件。SCI。 技术。 25,062002(2012)。 [22] Matsumoto Kaname:应用物理77,19(2008)。 [23] Yamada Shigeru:应用物理93,206(2024)。 [24] Y. Yamada,第36届国际超导性国际研讨会(ISS2023),Takina,新西兰惠灵顿,11月28日至30日,2023年。 [25] Miyata Noboru:材料37,361(1988)。 [26] https://www.t.u-tokyo.ac.jp/press/pr2023-06-28-001 [27] A. Stangl等。 ,科学。 Rep。11,8176(2021)。 [28] R. Hiwatari等。 ,血浆融合res。 14,1305047(2019)。 [29]在美国休斯顿大学申请2023年国际申请指挥的布兰登·索博姆(Brandon Sorbom)(2023年)。 [30] D. uglietti,超越。 SCI。 技术。 32,053001(2019)。SCI。技术。25,062002(2012)。[22] Matsumoto Kaname:应用物理77,19(2008)。[23] Yamada Shigeru:应用物理93,206(2024)。[24] Y. Yamada,第36届国际超导性国际研讨会(ISS2023),Takina,新西兰惠灵顿,11月28日至30日,2023年。[25] Miyata Noboru:材料37,361(1988)。[26] https://www.t.u-tokyo.ac.jp/press/pr2023-06-28-001 [27] A. Stangl等。,科学。Rep。11,8176(2021)。 [28] R. Hiwatari等。 ,血浆融合res。 14,1305047(2019)。 [29]在美国休斯顿大学申请2023年国际申请指挥的布兰登·索博姆(Brandon Sorbom)(2023年)。 [30] D. uglietti,超越。 SCI。 技术。 32,053001(2019)。Rep。11,8176(2021)。[28] R. Hiwatari等。,血浆融合res。14,1305047(2019)。[29]在美国休斯顿大学申请2023年国际申请指挥的布兰登·索博姆(Brandon Sorbom)(2023年)。[30] D. uglietti,超越。SCI。 技术。 32,053001(2019)。SCI。技术。32,053001(2019)。

在Caudoputamen和Accumbens核中直接和间接途径的纹状体投影神经元中树突状刺的差异发育
产后发育中的突触修饰对于神经网络的成熟至关重要。兴奋性突触的发育成熟发生在树突状棘的基因座,受生长和修剪动态调节。纹状体棘投射神经元(SPN)从大脑皮层和thalaus中获得兴奋性输入。spns和纹状体层间间接途径(ISPN)的SPN具有不同的发育根和功能。这两种类型的SPN的树突状脊柱成熟的时空动力学仍然难以捉摸。在这里,我们描绘了伏齿木剂和伏齿核(NAC)中DSPN和ISPN的树突状刺的发育轨迹。我们通过将Cre依赖性的AAV-EYFP病毒微注射到新生儿DRD1-CRE或Adora2a-Cre小鼠中,并通过微注射CRE依赖性AAV-EYFP病毒标记了SPN的树突状刺,并在三个级别上分析了旋转生成,包括不同的SPN细胞类型,子区域和后期。在背外侧纹状体中,DSPN和ISPN的脊柱修剪发生在产后(P)30 - P50。在背侧纹状体中,DSPN和ISPN的脊柱密度在P30和P50之间达到了峰值,而DSPN和ISPN的脊柱修剪分别发生在P30和P50之后。在NAC壳中,在p21 - P30后修剪DSPN和ISPN的棘突,但在NAC外侧壳的ISPN中未观察到明显的修剪。在NAC核心中,DSPN和ISPN的脊柱密度分别达到P21和P30的峰值,随后下降。总体而言,DSPN和ISPN中树突状棘的发育成熟遵循背侧和腹侧纹状体中不同的海上轨迹。


植物病原糸状菌ゲノム编集技术の开発†
† 令和 2 年 3 月 19 日 令和 2 年度大会で行われる予定であった学术奨励赏研究の目的 * 东京理科大学理工学部応用生物科学科 Department of Applied Biological Science, Professor of Science and Technology, Tokyo University of Science, 2641 Yamazaki, Noda-shi, Chiba 278-8510,日本

发育中的隔膜中神经元的独特“逆向”迁移
屏状核(CLA)是位于岛叶皮质和纹状体之间的一簇神经元。许多研究表明,CLA 在高级大脑功能中起着重要作用。此外,越来越多的证据表明 CLA 功能障碍与神经心理症状有关。然而,CLA 在发育过程中是如何形成的尚不完全清楚。在本研究中,我们分析了 CLA 的发育,特别关注了雌雄小鼠中 CLA 神经元的迁移情况。首先,我们发现 CLA 神经元是在胚胎第 10.5 天和第 12.5 天之间产生的,但大部分是在第 11.5 天产生的。接下来,我们使用 FlashTag 技术标记了在 E11.5 出生的 CLA 神经元,并发现大多数神经元在 E13.5 时到达大脑表面,但在 1 天后的 E14.5 时分布在 CLA 深处。GFP 标记细胞的延时成像显示,一些 CLA 神经元首先向外径向迁移,然后在到达表面后改变方向向内迁移。此外,我们证明了 Reelin 信号对于 CLA 神经元的适当分布是必需的。发育中的 CLA 神经元从向外迁移到“反向”迁移的转变与其他迁移模式不同,在其他迁移模式中,神经元通常沿某个方向迁移,即简单的向外或向内。未来对 CLA 发育特征和精确分子机制的阐明可能会为 CLA 独特的认知功能提供见解。
基因编辑
将打靶特定人源基因的 Cas9 和 sgRNA 转染到 HEK293 细胞。转染所用的质粒 DNA 上含有 表达带双端核定位序列 ( NLS )的 Cas9 及 sgRNA 的表达框,通过 TransIT-X2 (Mirus) 转染 试剂进行转染。转染所用的 Cas9 mRNA 进行了假尿苷和 5- 甲基胞嘧啶修饰且带有双端 核定位序列,使用 transIT-mRNA 转染试剂将 sgRNA 和 mRNA 共转染。 Cas9 RNPs 使用脂质 体 RNAiMAX ( Life Technologies ) 进行反向转染, RNP 的终浓度为 10 nmol 。 Cas9 蛋白上不含 核定位序列。 EnGen Cas9 含有双端核定位序列。编辑效率通过 T7E1 实验进行分析,结果 以修饰百分比进行统计。