XiaoMi-AI文件搜索系统
World File Search System异构体

植物SUMO化对抗病毒的两面性
与大多数生物体一样,植物也具备复杂而精巧的分子机制来应对不断变化的环境。在翻译后修饰 (PTM) 中,小肽(如泛素或 SUMO(小泛素相关修饰物))的结合能够快速有效地适应各种非生物和生物胁迫条件。SUMO 化过程涉及使用类似于泛素化的分级多酶级联将 SUMO 共价附着到目标蛋白上(图 1)[ 1 ]。这种可逆修饰可导致构象变化、改变蛋白质相互作用并影响修饰蛋白质的整体功能,包括稳定性、亚细胞定位和转录调控。除了与目标蛋白结合之外,SUMO 还能够与许多含有 SUMO 相互作用基序 (SIM) 的蛋白质非共价相互作用。将相同或不同蛋白质中的 SUMO 化位点与 SIM 相结合,有助于形成蛋白质宏观结构,从而通过将其他 SUMO 靶标募集到有利于 SUMO 化的环境中来增强 SUMO 化 [1]。拟南芥基因组含有 8 个 SUMO 基因,但只有 4 个得到表达(AtSUMO1/2/3/5)。几乎相同的 AtSUMO1/2 是 SUMO 原型,因为它们是哺乳动物 SUMO2/3 的最近同源物。SUMO 蛋白在发育和防御过程中的时空表达和功能有所不同 [2]。植物通常表达高水平的高度保守的 SUMO 异构体(AtSUMO1/2)和至少一种弱表达的非保守异构体(AtSUMO3/5)。

人类 DHFR2 基因的差异翻译能力表明存在低丰度的发育和组织特异性内源性蛋白质
根据预测翻译蛋白的重组版本的酶活性,人类二氢叶酸还原酶 2 ( DHFR2 ) 基因已被归因于功能性作用。然而,其体内功能仍不清楚。DHFR2 与其亲本同源物 DHFR 之间的高氨基酸序列同一性 (92%) 使内源性蛋白质的分析具有挑战性。本文介绍了一种针对几种人类细胞系和组织类型的靶向质谱蛋白质组学方法,以识别 DHFR2 特异性肽作为其翻译的证据。我们提供了确凿的证据,表明线粒体中的 DHFR2 活性实际上是由 DHFR 而不是 DHFR2 介导的。 Ribo-seq 数据分析和使用蔗糖垫进行的核糖体关联实验评估表明,Ensembl 注释的 DHFR2 的两个主要 mRNA 异构体 201 和 202 与核糖体存在不同的关联。这表明它在 RNA 和蛋白质水平上都发挥着功能性作用。然而,尽管 DHFR2 的各种 RNA 异构体相对丰富,但我们无法在大多数细胞类型中检测到可检测水平的 DHFR2 蛋白。我们确实在胚胎心脏中检测到了 DHFR2 特异性肽,这表明该蛋白质可能在胚胎发生过程中发挥特殊作用。我们认为 DHFR2 基因在成体细胞中的主要功能很可能出现在 RNA 水平上。

全球群体基因组学揭示关键海洋植物线粒体基因组结构的长期稳定性
花植物的线粒体基因组 (mitogenome) 由多条染色体组成。线粒体染色体内和染色体之间的重组可能产生称为异构体的多种 DNA 分子。由于不均匀的复制和同源重组,异构体的拷贝数和组成在单个植物内和单个植物之间可能是动态的。尽管如此,尽管它们具有功能重要性,但物种内线粒体基因组的保守水平仍未得到充分研究。个体发育变异是否会导致线粒体基因组组成的世代进化目前尚不清楚。在这里,我们表明,海草 Zostera marina 的线粒体基因组组成在大约 35 万年前分化的全球种群中是保守的。使用长读测序,我们表征了 Z. marina 线粒体基因组并推断出重组诱导配置的库。为了描述全球线粒体基因组结构并研究其进化,我们研究了从太平洋和大西洋的 16 个种群中取样的 Z. marina 分生区域的线粒体基因组。我们的研究结果显示,同工型相对拷贝数具有惊人的相似性,这表明尽管在个体发育过程中存在显著的变化,但远亲种群和植物种系中的线粒体基因组组成具有高度的保守性。我们的研究为在植物个体水平上对动态线粒体基因组的观察与长期线粒体进化之间提供了联系。

高级难度的字段
高级难度理论的领域1。立体化学纽曼预测;控制新的立体中心(Felkin-Anh,Zimmerman-Traxler)的模型;方形平面和八面体过渡金属复合物的几何异构体;识别具有多个立体中心的分子中的异构体可能性。2。酶根据反应类型分类;同位素标记研究;涉及辅酶A的代谢途径A。3。相位和化学平衡潜热和Clausius-Clapeyron方程;综合性能;平衡常数的温度依赖性。4。分析技术质谱法(分子离子,碎片,同位素分布); IR数据的解释。5。光化学光催化;乐队间隙;量子产量;半导体。6。mo理论mo图的硅藻图;金属 - 配体相互作用。The following topics will not appear at IChO 2025: Formal group theory Planar, axial, or helical chirality Enzymatic kinetics Quantitative understanding of any isotope effects Kinetics of complex reactions Steady state and quasi equilibrium approximations NMR spectroscopy Synthetic polymers Photocatalytic organic mechanisms Pericyclic organic mechanisms Crystal field theory Thermodynamics and kinetics of吸附固态晶体结构不预期:记住心脏实用的代谢途径1。真空过滤2。薄层色谱图3。微观底片和96井板的使用显微镜反应不会出现在ICHO 2025上:不预期使用不混可能的溶剂来提取学生的提取:使用:使用分光光度计本身

EECS 280计算机视觉春季2025 Jitendra Malik,Angjoo KanazawaEECS 280计算机视觉春季2025 Jitendra Malik,Angjoo Kanazawa
要使这些概念更加精确,我们需要发展欧几里得转型的基本理论。一组转换定义了“一致性”或具有相同形状的概念。在高中的几何形状中,我们了解到两个平面三角形是一致的,如果其中一个可以旋转和翻译,以便恰好位于另一个平面。旋转和翻译是欧几里得转化的例子,也称为异构体或刚体运动,定义为保留任何一对点之间距离的变换。当我移动椅子时,这在椅子上的任何一对点之间都保持真实,但显然不是在气球上膨胀的点上。

ADAR1 编辑组揭示了编辑特异性的驱动因素......
作用于 RNA ADAR1 的腺苷脱氨酶促进双链和结构化 RNA 中的 A 到 I 转换。ADAR1 有两种异构体,它们从不同的启动子转录:细胞质 ADAR1p150 是干扰素诱导的,而 ADAR1p110 是组成性表达的,主要位于细胞核中。ADAR1 突变会导致艾卡迪-戈蒂埃综合征 (AGS),这是一种与异常 IFN 产生相关的严重自身炎症疾病。在小鼠中,ADAR1 或 p150 异构体的缺失会导致胚胎死亡,这是由干扰素刺激基因的过度表达引起的。这种表型通过删除细胞质 dsRNA 传感器 MDA5 得到挽救,表明 p150 同工型是不可或缺的,不能被 ADAR1p110 挽救。尽管如此,ADAR1p150 唯一针对的编辑位点仍然难以捉摸。在这里,通过将 ADAR1 同工型转染到无 ADAR 的小鼠细胞中,我们检测到了同工型特异性的编辑模式。使用突变的 ADAR 变体,我们测试了细胞内定位和 Z-DNA 结合域的存在如何影响编辑偏好。这些数据表明 ZBD 对 p150 编辑特异性的贡献很小,而同工型特异性编辑主要由 ADAR1 同工型的细胞内定位指导。我们的研究通过对异位表达标记 ADAR1 同工型的人类细胞的 RIP-seq 进行补充。两个数据集均表明 ADAR1p110 富集了内含子编辑和结合,而 ADAR1p150 优先结合和编辑 3'UTR。
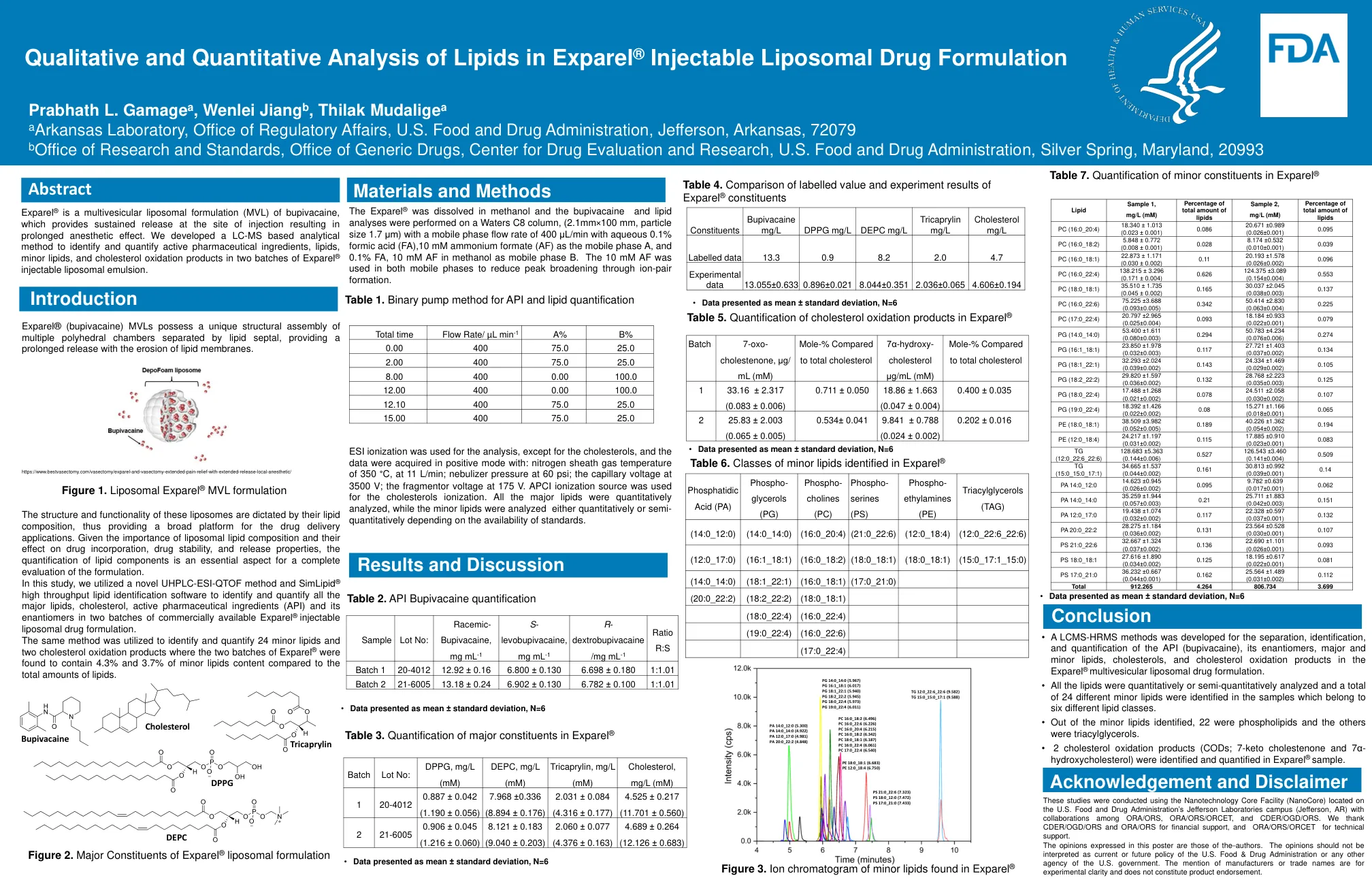
exparel中脂质的定性和定量分析
•开发了一种LCMS-HRMS方法,用于对API(Butivacaine)的分离,鉴定和定量,其对映异构体,主要脂质,胆固醇和胆固醇和胆固醇氧化产物在Exparel®多蛋白脂质体药物制剂中。•对所有脂质进行了定量或半定量分析,并在属于六个不同脂质类别的样品中鉴定了24个不同的小脂质。•从确定的小脂质中,有22个是磷脂,其他是三酰基甘油。•2种胆固醇氧化产物(COD; 7-酮胆固醇和7α-羟基胆固醇)在Exparel®样品中鉴定并量化。

最终意见 19/24
领域背景和常识:肌醇是作为立体异构体存在的糖醇,它们具有相似的化学结构,但空间取向不同。在已知的九种立体异构体中,MYO 和 DCI 在自然界中最为常见。人体从葡萄糖合成 MYO,并通过酶差向异构酶 (O1) 将部分 MYO 转化为 DCI。自然界中,DCI 存在于角豆荚和某些豆类中,而 MYO 存在于柑橘类水果和特定豆类中 (O2、O3)。MYO 和 DCI 都在胰岛素信号通路中发挥关键作用。它们的缺乏与胰岛素抵抗有关,胰岛素抵抗是一种身体对胰岛素反应不当的疾病,导致高血糖症 (O4、O5) 等代谢问题。胰岛素抵抗是多囊卵巢综合征 (PCOS) 和糖尿病 (O5) 等疾病的标志。 PCOS 是一种常见的激素紊乱,其特征是代谢功能障碍,包括高血糖、胆固醇水平异常、高血压和胰岛素抵抗 (O5)。这种情况通常会导致雄激素水平升高(睾酮等男性激素)、生育问题和月经周期不规律。由于胰岛素抵抗 (O6),PCOS 患者体重增加也很常见。在专利申请时,已经充分证实 MYO 和 DCI 补充剂可以改善胰岛素抵抗并缓解 PCOS 患者的症状 (O7、O8)。这两种肌醇都被认为是安全的,可供人类食用,但专利范围不仅限于人类使用,还扩展到任何潜在应用。这些信息被视为本领域的专业人士的常识,因为有多项研究和临床观察支持。

EXO70 和 MLO 蛋白的相互作用调节毛状体细胞壁组成和对白粉病的易感性
70 kDa (EXO70) 蛋白的胞外囊泡成分是胞外囊泡复合物的组成部分,与胞吐过程中的囊泡束缚有关。抗霉菌位点 O (MLO) 蛋白是植物特异性钙通道,一些 MLO 同工型可促进真菌白粉病的致病。我们在此检测到拟南芥 exo70H4 和 mlo2 mlo6 mlo12 三重突变体植物在叶毛状体次生细胞壁的生物发生方面存在意外的表型重叠。生化和傅里叶变换红外光谱分析证实了这些突变体中毛状体细胞壁组成的缺陷。表达荧光团标记的 EXO70H4 和 MLO 的转基因系表现出这些蛋白质的广泛共定位。此外,mCherry-EXO70H4 错误定位在 mlo 三重突变体的毛状体中,反之亦然,MLO6-GFP 错误定位在 exo70H4 突变体的毛状体中。GFP 标记的 PMR4 胼胝体合酶(EXO70H4 依赖性胞吐的已知货物)的表达表明,mlo 三重突变体植物的毛状体中 GFP-PMR4 的细胞壁输送减少。植物和酵母细胞中的体内蛋白质-蛋白质相互作用测定揭示了 EXO70.2 亚家族成员和 MLO 蛋白之间的异构体优先相互作用。最后,exo70H4 和 mlo6 突变体结合时表现出协同增强的对白粉病攻击的抗性。总之,我们的数据表明 EXO70 和 MLO 蛋白在调节毛状体细胞壁生物合成和白粉病易感性方面存在异构体特异性相互作用。

控制缺氧诱导因子靶标搜索和结合动力学的机制
摘要 转录因子 (TF) 通常被认为是一种模块化结构,包含结构良好的序列特异性 DNA 结合结构域 (DBD) 与无序的激活结构域 (AD) 配对,后者负责靶向辅助因子或核心转录起始机制的蛋白质-蛋白质相互作用。然而,这种简单的分工模型无法解释为什么在体外确定的具有相同 DNA 结合序列特异性的 TF 在体内表现出不同的结合谱。缺氧诱导因子 (HIF) 家族提供了一个鲜明的例子:在几种癌症类型中异常表达的 HIF-1 α 和 HIF-2 α 亚基异构体在体外识别相同的 DNA 基序——缺氧反应元件 (HRE)——但在体内仅共享其靶基因的一个子集,同时在某些情况下对癌症的发展和进展产生对比的影响。为了探究介导异构体特异性基因调控的机制,我们使用活细胞单粒子追踪 (SPT) 来研究 HIF 核动力学及其在遗传扰动或药物治疗下的变化。我们发现 HIF-α 亚基及其二聚化伴侣 HIF-1β 表现出独特的扩散和结合特性,这些特性对浓度和亚基化学计量极为敏感。使用域交换变体、突变和 HIF-2α 特异性抑制剂,我们发现尽管 DBD 和二聚化域很重要,但染色质结合和扩散行为的另一个主要决定因素是含有 AD 的内在无序区域 (IDR)。使用 Cut&Run 和 RNA-seq 作为正交基因组方法,我们还证实了 IDR 依赖的 HIF 靶基因特定子集的结合和激活。这些发现揭示了 IDR 在调节 TF 搜索和结合过程中以前未被重视的作用,这有助于染色质上的功能性靶位点选择性。