XiaoMi-AI文件搜索系统
World File Search System新霉素
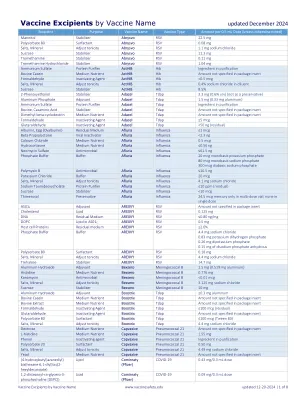
疫苗辅料(按疫苗名称)
甘露醇稳定剂 Abrysvo RSV 22.5 mg 聚山梨醇酯 80 表面活性剂 Abrysvo RSV 0.08 mg 矿物质盐 调节张力 Abrysvo RSV 1.1 mg 氯化钠 蔗糖稳定剂 Abrysvo RSV 11.3 mg 氨丁三醇稳定剂 Abrysvo RSV 0.11 mg 盐酸氨丁三醇稳定剂 Abrysvo RSV 1.04 mg 硫酸铵 蛋白质净化剂 ActHIB Hib 净化成分 牛酪蛋白培养基营养物 ActHIB Hib 包装说明书中未指定的数量 甲醛灭活剂 ActHIB Hib <0.5 mcg 矿物质盐 调节张力 ActHIB Hib 稀释剂中 0.4% 氯化钠 蔗糖稳定剂 ActHIB Hib 8.5% 2-苯氧乙醇稳定剂 Adacel Tdap 3.3 mg (0.6% v/v)(不作为防腐剂) 磷酸铝佐剂 Adacel Tdap 1.5 mg(0.33 mg 铝) 硫酸铵 蛋白质净化剂 Adacel Tdap 净化成分 牛,酪蛋白氨基酸稳定剂 Adacel Tdap 包装说明书中未指定的数量 二甲基-β-环糊精 培养基营养物 Adacel Tdap 包装说明书中未指定的数量 甲醛灭活剂 Adacel Tdap ≤5 mcg 戊二醛灭活剂 Adacel Tdap <50 ng(残留) 蛋清(卵清蛋白) 残留培养基 Afluria Influenza ≤1 mcg β-丙内酯病毒灭活剂 Afluria Influenza <2.3 ng 氯化钙 培养基营养物 Afluria Influenza 0.5 mcg 氢化可的松 培养基营养物 Afluria Influenza ≤0.56 ng 硫酸新霉素 抗菌剂 Afluria Influenza ≤61.5 ng 磷酸盐缓冲液 缓冲液 Afluria Influenza 20 mcg 磷酸二氢钾 80 mcg 磷酸二氢钠 300 mcg 磷酸二氢钠 多粘菌素 B 抗菌剂 Afluria Influenza ≤10.5 ng 氯化钾缓冲液 Afluria Influenza 20 mcg 矿物质盐 调节张力 Afluria Influenza 4.1 mg 氯化钠 牛磺脱氧胆酸钠 蛋白质净化剂 Afluria Influenza ≤10 ppm(残留) 蔗糖稳定剂 Afluria Influenza <10 mcg 硫柳汞防腐剂 Afluria Influenza 24.5 mcg 汞仅在多剂量小瓶中;单剂量中无 AS01 E 佐剂 AREXVY RSV 包装说明书中未指定的量 胆固醇 脂质 AREXVY RSV 0.125 mg DNA 残留培养基 AREXVY RSV ≤0.80 ng/mg DOPC AS01 E 中的脂质 AREXVY RSV 0.5 mg 宿主细胞 蛋白质 残留培养基 AREXVY RSV ≤2.0% 磷酸盐缓冲液 缓冲液 AREXVY RSV 4.4 mg 氯化钠 0.83 mg 磷酸二氢钾 0.26 mg 磷酸二钾 0.15 mg 无水磷酸二钠 聚山梨醇酯 80 表面活性剂 AREXVY RSV 0.18 mg 盐、矿物质 调节张力 AREXVY RSV 4.4 mg 氯化钠 海藻糖 稳定剂 AREXVY RSV 14.7 mg 氢氧化铝 佐剂 Bexsero 脑膜炎球菌B 1.5 毫克(0.519 毫克铝)组氨酸培养基营养素Bexsero 脑膜炎球菌 B 0.776 毫克卡那霉素抗菌素Bexsero 脑膜炎球菌 B <0.01 微克盐、矿物质调节张力Bexsero 脑膜炎球菌 B 3.125 毫克氯化钠蔗糖稳定剂Bexsero 脑膜炎球菌 B 10 毫克氢氧化铝佐剂增强剂Tdap ≤0.3 毫克铝 牛酪蛋白 培养基 营养物 Boostrix Tdap 包装说明书中未指定量 牛提取物 培养基 营养物 Boostrix Tdap 包装说明书中未指定量 甲醛灭活剂 Boostrix Tdap ≤100 微克(残留) 戊二醛灭活剂 Boostrix Tdap 包装说明书中未指定量 聚山梨醇酯 80 表面活性剂 Boostrix Tdap ≤100 微克(吐温 80) 矿物质盐 调节张力 Boostrix Tdap 4.4 毫克氯化钠 葡萄糖 培养基 营养物 Capvaxive 肺炎球菌 21 包装说明书中未指定量 L-组氨酸 培养基 营养物 Capvaxive 肺炎球菌 21 1.55 毫克 苯酚灭活剂 Capvaxive 肺炎球菌 21 净化成分 聚山梨醇酯 20 表面活性剂Capvaxive 肺炎球菌 21 0.50 mg 盐、矿物质 调节张力 Capvaxive 肺炎球菌 21 4.49 mg 氯化钠 酵母培养基营养物 Capvaxive 肺炎球菌 21 包装说明书中未指定的量 (4-羟基丁基)氮烷二基)双(己烷-6,1-二基)双(2-己基癸酸酯)
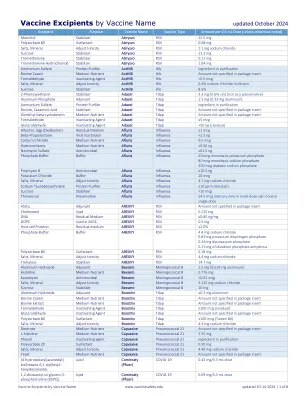
疫苗辅料(按疫苗名称)
甘露醇稳定剂 Abrysvo RSV 22.5 mg 聚山梨醇酯 80 表面活性剂 Abrysvo RSV 0.08 mg 矿物质盐 调节张力 Abrysvo RSV 1.1 mg 氯化钠 蔗糖稳定剂 Abrysvo RSV 11.3 mg 氨丁三醇稳定剂 Abrysvo RSV 0.11 mg 盐酸氨丁三醇稳定剂 Abrysvo RSV 1.04 mg 硫酸铵 蛋白质净化剂 ActHIB Hib 净化成分 牛酪蛋白培养基营养物 ActHIB Hib 包装说明书中未指定的数量 甲醛灭活剂 ActHIB Hib <0.5 mcg 矿物质盐 调节张力 ActHIB Hib 稀释剂中 0.4% 氯化钠 蔗糖稳定剂 ActHIB Hib 8.5% 2-苯氧乙醇稳定剂 Adacel Tdap 3.3 mg (0.6% v/v)(不作为防腐剂) 磷酸铝佐剂 Adacel Tdap 1.5 mg(0.33 mg 铝) 硫酸铵 蛋白质净化剂 Adacel Tdap 净化成分 牛,酪蛋白氨基酸稳定剂 Adacel Tdap 包装说明书中未指定的数量 二甲基-β-环糊精 培养基营养物 Adacel Tdap 包装说明书中未指定的数量 甲醛灭活剂 Adacel Tdap ≤5 mcg 戊二醛灭活剂 Adacel Tdap <50 ng(残留) 蛋清(卵清蛋白) 残留培养基 Afluria Influenza ≤1 mcg β-丙内酯病毒灭活剂 Afluria Influenza <2.3 ng 氯化钙 培养基营养物 Afluria Influenza 0.5 mcg 氢化可的松 培养基营养物 Afluria Influenza ≤0.56 ng 硫酸新霉素 抗菌剂 Afluria Influenza ≤61.5 ng 磷酸盐缓冲液 缓冲液 Afluria Influenza 20 mcg 磷酸二氢钾 80 mcg 磷酸二氢钠 300 mcg 磷酸二氢钠 多粘菌素 B 抗菌剂 Afluria Influenza ≤10.5 ng 氯化钾缓冲液 Afluria Influenza 20 mcg 矿物质盐 调节张力 Afluria Influenza 4.1 mg 氯化钠 牛磺脱氧胆酸钠 蛋白质净化剂 Afluria Influenza ≤10 ppm(残留) 蔗糖稳定剂 Afluria Influenza <10 mcg 硫柳汞防腐剂 Afluria Influenza 24.5 mcg 汞仅在多剂量小瓶中;单剂量中无 AS01 E 佐剂 AREXVY RSV 包装说明书中未指定的量 胆固醇 脂质 AREXVY RSV 0.125 mg DNA 残留培养基 AREXVY RSV ≤0.80 ng/mg DOPC AS01 E 中的脂质 AREXVY RSV 0.5 mg 宿主细胞 蛋白质 残留培养基 AREXVY RSV ≤2.0% 磷酸盐缓冲液 缓冲液 AREXVY RSV 4.4 mg 氯化钠 0.83 mg 磷酸二氢钾 0.26 mg 磷酸二钾 0.15 mg 无水磷酸二钠 聚山梨醇酯 80 表面活性剂 AREXVY RSV 0.18 mg 盐、矿物质 调节张力 AREXVY RSV 4.4 mg 氯化钠 海藻糖 稳定剂 AREXVY RSV 14.7 mg 氢氧化铝 佐剂 Bexsero 脑膜炎球菌B 1.5 毫克(0.519 毫克铝)组氨酸培养基营养素Bexsero 脑膜炎球菌 B 0.776 毫克卡那霉素抗菌素Bexsero 脑膜炎球菌 B <0.01 微克盐、矿物质调节张力Bexsero 脑膜炎球菌 B 3.125 毫克氯化钠蔗糖稳定剂Bexsero 脑膜炎球菌 B 10 毫克氢氧化铝佐剂增强剂Tdap ≤0.3 毫克铝 牛酪蛋白 培养基 营养物 Boostrix Tdap 包装说明书中未指定量 牛提取物 培养基 营养物 Boostrix Tdap 包装说明书中未指定量 甲醛灭活剂 Boostrix Tdap ≤100 微克(残留) 戊二醛灭活剂 Boostrix Tdap 包装说明书中未指定量 聚山梨醇酯 80 表面活性剂 Boostrix Tdap ≤100 微克(吐温 80) 矿物质盐 调节张力 Boostrix Tdap 4.4 毫克氯化钠 葡萄糖 培养基 营养物 Capvaxive 肺炎球菌 21 包装说明书中未指定量 L-组氨酸 培养基 营养物 Capvaxive 肺炎球菌 21 1.55 毫克 苯酚灭活剂 Capvaxive 肺炎球菌 21 净化成分 聚山梨醇酯 20 表面活性剂Capvaxive 肺炎球菌 21 0.50 mg 盐、矿物质 调节张力 Capvaxive 肺炎球菌 21 4.49 mg 氯化钠 酵母培养基营养物 Capvaxive 肺炎球菌 21 包装说明书中未指定的量 (4-羟基丁基)氮烷二基)双(己烷-6,1-二基)双(2-己基癸酸酯)

生物学Pogil蛋白合成的中心教条
弗朗西斯·克里克(Francis Crick)的分子生物学基本原理包括序列假设和中央教条。序列假说描述了核酸和蛋白质之间如何转移序列信息。中央教条概述了遗传信息从DNA到RNA,然后概述了蛋白质,并指出一旦该信息达到蛋白质水平,就无法检索。近年来,合成生物学对这些原则提出了挑战,提出了有关潜在违反中心教条的问题。为了解决这些问题,研究人员在蛋白质合成中的信息传递和CRISPR基因编辑之间取得了相似之处。将三部分评估方案应用于CRISPR/CAS9和CRISPR Prime编辑系统。虽然信息传递保持在中央教条的范围内,但潜在的机制表明,通过合成生物学违反了这一原理的潜在途径。这引发了人们对蛋白质衍生的信息转移系统的理论和实际意义的猜测。此外,还有一项为入门生物学学生设计的教育活动,该活动使用诸如乙烯基记录等非生物学示例来探索中心教条。学生检查真核细胞中的遗传信息流,并探索逆转录病毒感染和伤口愈合等生物学条件,从而改变了这种流动。这个主题对于对医学,医疗保健和生物医学研究感兴趣的学生至关重要,因为遗传信息流的变化可能导致疾病状态。但是,这种简化不会捕获其原始含义。分子生物学的中心教条围绕着生物系统中的遗传信息流动,通常总结为“ DNA生成RNA,RNA产生蛋白质”。弗朗西斯·克里克(Francis Crick)在1957年介绍了这一概念,在1970年重申了这一概念:“分子生物学的中心教条涉及顺序信息的详细残基传递。它指出,这种信息不能从蛋白质转移到蛋白质或核酸。”更受欢迎但不正确的版本是简单的DNA→RNA→蛋白质途径,归因于詹姆斯·沃森(James Watson)。这个两步过程与Crick的原始声明不同,该声明今天仍然有效。包含DNA,RNA和蛋白质的生物聚合物是线性聚合物,每个单体最多都连接到其他两个。他们的序列有效地编码信息,并在分子之间发生忠实的,确定性的转移。当DNA转录为RNA时,它的补充对与它。DNA代码A,G,T和C分别转移到RNA代码A,G,U和C上。蛋白质的编码是使用人类和哺乳动物的标准密码子表中的三组,称为密码子。但是,有些生命形式使用不同的翻译。信息传递的基本步骤是从DNA到DNA的复制,必须为后代细胞提供遗传物质。复杂的蛋白质组重新组合执行此复制过程。转录是将DNA信息复制到mRNA中的过程。!!!可以发生替代剪接,从而增加蛋白质多样性。酶,例如RNA聚合酶和转录因子,促进了真核细胞的过程,包括剪接和翻译进行处理。转录过程始于在前mRNA中添加5'帽和poly-a尾巴,然后进行剪接。成熟的mRNA然后向核糖体传播进行翻译。在原核生物中,转录和翻译是连接的,而在真核生物中,它们通过mRNA从核从核转移到细胞质的转运而分开。翻译涉及通过核糖体读取mRNA密码子,将氨基酰基的TRNA匹配到抗代码,并将氨基酸连接到生长的肽链中。链条开始折叠成正确的构象,最终蛋白质出现所需的其他处理。翻译以终止密码子结束,但是mRNA不包含指定成熟蛋白质所需的所有信息。处理对于正确折叠至关重要,通常涉及伴侣蛋白以及多肽链的剪接或分裂。某些蛋白质需要交联,辅因子附着或去除内部。新霉素可以增强从其他生物体分离的单链DNA模板中合成蛋白质。但是,目前尚不清楚这种翻译机制是否针对遗传密码。翻译蛋白氨基酸序列后,可以通过酶编辑它们,该过程未被中央教条明确覆盖。没有太多明确的例子,蛋白质修饰和遗传学的相关概念彼此之间有很大关系。例子包括一些抗生素。某些蛋白质是由非核糖体肽合成酶合成的,这些蛋白质可以是专门合成一种类型的肽的复杂蛋白质。这些肽通常具有环状和/或分支结构,并且可以含有非蛋白酶氨基酸,从而将其与核糖体合成蛋白区分开。inteins是蛋白质的“寄生”片段,可以从氨基酸链中从核糖体出来并用肽键重新加入其余部分。此过程改变了蛋白质的主要序列,使其可以直接编辑DNA序列并增加其可遗传的繁殖。表观遗传学是指可以显着改变基因表达水平的DNA甲基化状态的变化。当信息状态的变化不会因体细胞突变而改变时,这种表观遗传变化被认为是可遗传的。prion是具有特定氨基酸序列的蛋白质和构象,它们通过在蛋白质的其他蛋白质分子中以相同的序列但不同的构象进行构象变化来传播自身。一旦蛋白质转换,它就可以将信息传达到新的细胞中,并将该序列的更多功能分子重新配置为替代prion形式。一些科学家认为,prion介导的遗传违反了分子生物学的核心教条。prion假说与分子生物学的核心教条矛盾,该序列指出DNA序列编码蛋白质信息。詹姆斯·夏皮罗(James A.相反,它提出蛋白质可以包含其自己的遗传信息,影响其生物学功能,并可能被传递给其他分子。但是,他的批评家不相信他对克里克最初意图的解释。弗朗西斯·克里克(Francis Crick)在自传中写道,他选择“中央教条”一词,并指出他想强调其重要性和力量。后来他意识到,使用“教条”一词造成的麻烦远远超出了价值,因为它暗示了一种信念,这是无法怀疑的。Crick将该术语用作口号,但并未完全理解其正确的含义。中央教条的概念早于发现DNA的作用和结构。八月魏斯曼的种植血浆理论提出,遗传信息仅从种系细胞转向体细胞,预测了以基因为中心的生命观点,而无需直接解决分子生物学。分子生物学的中心教条,这是弗朗西斯·克里克(Francis Crick)在1950年代创造的概念,概述了遗传信息从DNA到蛋白质的流动。根据此框架,遗传信息存储在DNA分子中,并转录为RNA,然后将其转化为蛋白质。然而,随着研究人员继续探索细胞过程的复杂性,他们开始质疑中央教条的简单性。尤其是,prions的发现 - 可以改变基因表达的传染性蛋白质颗粒 - 挑战了遗传信息无法从蛋白质向后流向DNA的观念。在这种情况下,必须重新评估中心教条在现代生物学中的相关性至关重要。最近的研究进一步模糊了传统的中央教条与非编码RNA介导的基因调节的概念之间的界限。后者表明,非编码RNA可以通过充当分子“海绵”或直接与染色质重塑复合物相互作用来影响基因表达。一些科学家认为,应扩展中央教条,以包括非编码RNA的作用以及允许蛋白质影响基因表达的反馈机制。