XiaoMi-AI文件搜索系统
World File Search System无性繁殖

达卡大学植物学系教学大纲...达卡大学植物学系教学大纲...
完整分数:100 1。分类学中的解剖学证据:叶子解剖学;气孔,毛状体;叶柄解剖叶面叶子;茎解剖学;静脉解剖学。2。生殖生物学:人口增长,大小,密度和承载能力;在繁殖系统中,繁殖系统,无性繁殖,与系统学有关的生殖生物学;研究方法;授粉类型,机制,向量。3。天然产品化学在分类学和系统发育中的应用:在解决趋化问题问题中要遵循的步骤;一些用于趋化性相关性的天然产物;色谱 - 基础和定义;通过蛋白质分析的趋化性。4。栖息地;环境关系;生物,非生物因素;空间关系,时间关系,适应性特征。生命形式,侨民类型,忠诚,活力,叶子大小,发育反应,杂物。5。生物多样性与保护:生物多样性保护概论;世界保护策略,孟加拉国的保护策略;国际生物多样性公约;现场保护的方法;孟加拉国的生物多样性保护问题和行动计划; IUCN红色列表类别和威胁孟加拉国的植物。

植物生物技术 40(3): 211-218 (2023)
摘要 基因组编辑对于作物改良非常有用。利用农杆菌中的瞬时表达系统表达基因组编辑酶的方法,称为农杆菌诱变,是基因组编辑技术中用于改良包括马铃薯在内的无性繁殖作物优良品种的一种捷径。然而,用这种方法不能选择经过编辑的个体。再生促进基因的瞬时表达可以导致幼苗再生出芽,而大多数再生促进基因的组成性表达不会导致正常再生的芽。在这里,我们报告我们可以通过正向选择获得基因组编辑的马铃薯。这些再生芽是通过将再生促进基因与基因组编辑酶基因的瞬时表达相结合的方法获得的。此外,我们证实,用这种方法获得的基因组编辑马铃薯不含有农杆菌中使用的二元载体的序列。我们的数据已提交给日本监管机构文部科学省 (MEXT),我们正在对这些马铃薯进行田间试验以进一步研究。我们的工作为通过再生促进基因的瞬时表达来再生和获取基因组编辑作物提供了一种强有力的方法。

12 种桉树的再生和农杆菌介导的遗传转化
摘要 桉树属有 900 多个品种和杂交种,其中许多是珍贵的速生硬木。由于其经济重要性,桉树是较早被破译基因组的树种之一。然而,缺乏有效的遗传转化系统严重制约了该植物的功能基因组学研究。桉树再生和转化的成功在很大程度上取决于基因型和外植体。在本研究中,我们系统地筛选了 12 个桉树品种的 26 个基因型,试图获得具有高再生潜力的桉树基因型。我们开发了两种常见的再生培养基,可用于大多数受试桉树基因型的播种下胚轴和克隆的节间作为外植体。然后,我们使用 DsRed2 作为遗传转化效率测试的视觉标记。我们的结果表明,E. camaldulen 和 E. robusta 适合进行遗传转化。最后,我们分别使用播种下胚轴和克隆节间成功地建立了稳定的农杆菌介导的桉树和桉树的遗传转化程序。总之,我们的研究为桉树的无性繁殖、基因转化、基于 CRISPR 的基因诱变、激活和抑制以及基因的功能表征提供了有价值的手段。

多个水平的微型染色体转移驱动器基因组的基因组演变
作物疾病大流行通常是由无性繁殖的植物病原体的克隆谱系驱动的。尽管遗传变异有限,并且在没有性重组的情况下,这些克隆病原体如何不断地适应其宿主。在这里,我们揭示了在爆炸真菌斑点的大流行克隆谱系中的水平染色体转移的多个实例(Syn。pyricularia)oryzae。我们确定了一个Hori Zontly转移的1.2MB辅助迷你染色体,该小染色体在大米爆炸真菌谱系和谱系感染印度鹅(Eleusine Indiona)的Oryzae分离株之间非常保守,这是一种经常生长的野生草,在耕种陶瓷毛皮的附近生长。此外,我们表明,这种迷你染色体是通过克隆大米爆炸株通过至少九个不同的转移事件水平获取的。这些发现建立了水平的迷你染色体转移,作为促进不同宿主相关的爆炸真菌谱系中遗传交换的一种机制。我们提出,感染野草的爆炸真菌是遗传储层,这些储层驱动了困扰谷物作物的大流行克隆谱系的基因组进化。

多个水平的微型染色体转移驱动器基因组的基因组演变
作物疾病大流行通常是由无性繁殖的植物病原体的克隆谱系驱动的。尽管遗传变异有限,并且在没有性重组的情况下,这些克隆病原体如何不断地适应其宿主。在这里,我们揭示了在爆炸真菌斑点的大流行克隆谱系中的水平染色体转移的多个实例(Syn。pyricularia)oryzae。我们确定了一个Hori Zontly转移的1.2MB辅助迷你染色体,该小染色体在大米爆炸真菌谱系和谱系感染印度鹅(Eleusine Indiona)的Oryzae分离株之间非常保守,这是一种经常生长的野生草,在耕种陶瓷毛皮的附近生长。此外,我们表明,这种迷你染色体是通过克隆大米爆炸株通过至少九个不同的转移事件水平获取的。这些发现建立了水平的迷你染色体转移,作为促进不同宿主相关的爆炸真菌谱系中遗传交换的一种机制。我们提出,感染野草的爆炸真菌是遗传储层,这些储层驱动了困扰谷物作物的大流行克隆谱系的基因组进化。

多个水平的微型染色体转移驱动器基因组的基因组演变
作物疾病大流行通常是由无性繁殖的植物病原体的克隆谱系驱动的。尽管遗传变异有限,并且在没有性重组的情况下,这些克隆病原体如何不断地适应其宿主。在这里,我们揭示了在爆炸真菌斑点的大流行克隆谱系中的水平染色体转移的多个实例(Syn。pyricularia)oryzae。我们确定了一个Hori Zontly转移的1.2MB辅助迷你染色体,该小染色体在大米爆炸真菌谱系和谱系感染印度鹅(Eleusine Indiona)的Oryzae分离株之间非常保守,这是一种经常生长的野生草,在耕种陶瓷毛皮的附近生长。此外,我们表明,这种迷你染色体是通过克隆大米爆炸株通过至少九个不同的转移事件水平获取的。这些发现建立了水平的迷你染色体转移,作为促进不同宿主相关的爆炸真菌谱系中遗传交换的一种机制。我们提出,感染野草的爆炸真菌是遗传储层,这些储层驱动了困扰谷物作物的大流行克隆谱系的基因组进化。

基于共编辑1策略2的T0代植物高效无转基因基因组编辑
对 T0 代植物进行无转基因基因组编辑是人们非常希望实现的目标,但同时也极具挑战性,尤其是在多年生植物和无性繁殖植物中。在这里,我们研究了通过农杆菌介导的胞嘧啶碱基编辑器 (CBE)/gRNA-Cas12a/crRNA-GFP 在植物体内瞬时表达来生成无转基因基因编辑植物的共编辑策略。具体而言,12 CBE/gRNA 用于碱基编辑 ALS 基因,从而赋予对除草剂氯磺隆的抗性,作为 13 选择标记,该抗性对植物表型没有负面影响;Cas12a/crRNA 用于编辑感兴趣的基因;GFP 用于选择无转基因转化子。使用 15 这种方法,可以在 T0 代中高效地针对番茄、烟草、马铃薯和柑橘中的各种基因 16 (单个或多个) 生成无转基因基因组编辑植物。除草剂抗性转化体中靶基因的双等位基因/纯合无转基因突变率在 8% 至 50% 之间。全基因组测序进一步证实了编辑植物中无转基因和无脱靶突变。共编辑策略对于在 T0 代中产生无转基因、基因组编辑植物是有效的,因此是植物遗传改良的有力工具。

农作物的非转基因基因组编辑方法
基于 CRISPR/Cas 的基因组编辑技术有可能加快大规模作物育种计划。然而,坚硬的细胞壁限制了 CRISPR/Cas 成分进入植物细胞,从而降低了基因组编辑效率。已建立的方法,例如农杆菌介导或基因枪转化,已用于将含有 CRISPR 成分的基因盒整合到植物基因组中。这些方法虽然有效,但也存在几个问题,包括 1) 转化过程需要费力且耗时的组织培养和再生步骤;2) 许多作物物种和优良品种难以转化;3) 在无性繁殖或高度杂合的作物(如菠萝)中,转基因的分离要么困难要么不可能;4) 生产转基因第一代可能引起公众争议和繁重的政府监管。无转基因基因组编辑技术的发展可以解决许多与转基因方法相关的问题。无转基因基因组编辑已通过递送预组装的 CRISPR/Cas 核糖核蛋白实现,尽管其应用有限。使用病毒载体递送 CRISPR/Cas 成分最近已成为一种强有力的替代方法,但需要进一步探索。在这篇综述中,我们讨论了无转基因基因组编辑方法的不同策略、原理、应用和未来方向。

一种高效的基于原生质体的葡萄属植物基因组编辑方案
CRISPR-Cas 技术可以对植物基因组进行精确修改,有望彻底改变农业。这些技术依赖于将编辑组件递送到植物细胞中以及再生完全编辑的植物。在无性繁殖植物(例如葡萄)中,原生质体培养是生产非嵌合和无转基因的基因组编辑植物的最佳途径之一。然而,植物从原生质体再生能力较差,阻碍了其用于基因组编辑的实施。在这里,我们报告了一种从来自多个葡萄品种的原生质体再生植物的有效方案。通过将原生质体封装在海藻酸钙珠中并与饲养层培养物共培养,原生质体分裂形成愈伤组织菌落,再生成胚胎并最终再生为植物。该方案成功应用于酿酒葡萄和鲜食葡萄(Vitis vinifera)品种,以及葡萄砧木和葡萄树野生近缘种 Vitis arizonica。此外,通过用 CRISPR-质粒或核糖核蛋白 (RNP) 复合物转染原生质体,我们在三个品种和 V. arizonica 中再生了 VvPHYTOENE DESATURASE 基因经过编辑的白化植物。结果揭示了该平台在促进葡萄属物种基因组编辑方面的潜力。
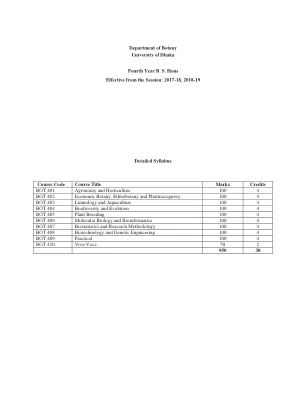
达卡大学植物学系四年级学士学位...
1. 引言:植物育种的定义、范围和目标。2. 作物的起源和驯化:作物起源中心、这一概念在植物育种中的重要性、作物的驯化。3. 植物遗传资源:种质的定义、收集、评估和保存(离体和就地),种质在植物育种计划中的应用。4. 生殖生物学和植物育种:作物的繁殖方式、植物改良中的授粉机制、自交不亲和性和雄性不育性及其在植物育种中的意义。5. 自花授粉和异花授粉作物的选择方法以及无性繁殖植物的克隆选择。6. 杂交:目标;杂交技术和类型以及人工杂交的重要性。7. 自花授粉和异花授粉作物的育种技术:大规模选择、纯系选择、谱系选择和批量方法的方法、优点和缺点。 8. 杂种优势育种:杂种优势、杂种优势和近交衰退的介绍,杂种优势的遗传基础,通过杂种优势育种取得的成果。9. 诱变育种:诱变育种介绍,植物诱变的人工诱导,诱变技术在作物改良中的应用,诱变育种的局限性。10. 回交育种:回交育种的方法、优点和局限性。11. 抗逆育种:逆境类型 - 生物和非生物,抗病育种方法,