XiaoMi-AI文件搜索系统
World File Search System有丝分裂

CLK 激酶抑制剂 1C8 和 GPS167 对上皮-间质转化和抗病毒免疫反应的影响揭示了它们的抗癌潜力
二杂芳酰胺基化合物 1C8 和氨基噻唑酰胺相关化合物 GPS167 可抑制 CLK 激酶,并影响多种癌细胞系的增殖。之前使用 GPS167 进行的化学基因组学筛选表明,与有丝分裂纺锤体组装相关的成分的消耗会改变对 GPS167 的敏感性。在这里,使用 1C8 进行的类似筛选也确定了参与有丝分裂纺锤体组装的成分的影响。因此,用 1C8 和 GPS167 处理的细胞的转录组分析表明,编码有丝分裂纺锤体组装成分的转录物的表达和 RNA 剪接受到影响。通过显示影响有丝分裂纺锤体组装的药物的亚毒性浓度会增加对 GPS167 的敏感性,证实了微管连接的功能相关性。1C8 和 GPS167 影响与肿瘤进展相关的通路中转录本的表达和剪接,包括 MYC 靶标和上皮间质转化 (EMT)。最后,1C8 和 GPS167 改变了参与抗病毒免疫反应的转录本的表达和可变剪接。与此观察结果一致,消耗双链 RNA 传感器 DHX33 可抑制 GPS167 介导的 HCT116 细胞细胞毒性。我们的研究揭示了 1C8 和 GPS167 影响癌细胞增殖以及转移关键过程的分子机制。

文章 - 人类和动物健康 Hec1/Nek2 有丝分裂通路抑制剂 INH1 抑制 A549 和 HeLa 细胞的细胞动力学参数
摘要:本研究在体外研究了 Hec1/Nek2 有丝分裂途径抑制剂 INH1 在腺癌人肺泡基底上皮细胞 A549 和人宫颈癌 HeLa 细胞系中的抗增殖作用。为此,使用了 xCELLigence 实时细胞分析 DP 仪器测定的细胞指数值、有丝分裂指数、BrdU 增殖测定和凋亡指数分析。本研究的结果表明,INH1 对 A549 具有细胞抑制和细胞骨架作用,对 HeLa 细胞具有细胞抑制作用。使用 xCelligence 设备测定两种细胞系的 IC 50 浓度均为 56 µM。所有其他参数均使用 IC 50 浓度。虽然该浓度降低了有丝分裂指数 BrdU 增殖值,但它增加了两种细胞系的凋亡指数值。对照组和实验组之间存在显着差异(p <0.05)。本研究的结果表明 INH1 可能成为不同类型癌症的有希望的治疗选择。

NDC80状态精确指标有丝分裂激酶抑制剂作为透明细胞肾细胞癌的新兴治疗选择
。cc-by-nc-nd 4.0国际许可证(未获得同行评审证书)获得的是作者/资助者,他已授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该版本的版权所有,该版本于2023年3月10日发布。 https://doi.org/10.1101/2023.03.07.531634 doi:biorxiv preprint

在有丝分裂神经元中持续表达UNC-4同源基因和unc-37/groucho,指定胆碱能突触的空间组织I
抽象的神经元细胞命运决定因素通过控制基因表达来调节神经元形态和突触连通性来确定神经元的身份。然而,尚不清楚神经元细胞命运决定因素是否具有突触模式形成的有丝分裂功能。在这里,我们在秀丽隐杆线虫的胆碱能运动神经元的瓷砖突触模式中确定了UNC-4同源蛋白及其Corepressor UNC-37/ Groucho的新作用。我们表明,在神经发生过程中不需要UNC-4,而是在有丝分裂后神经元中需要进行适当的突触模式。相比之下,在发育后和有丝分裂后神经元中都需要UNC-37。BAR-1/ B-蛋白突变抑制了UNC-4突变体的突触平铺缺陷,这对CEH-12/ HB9的表达进行了积极调节。异位CEH-12表达部分是UNC-4和UNC-37突变体的突触缺陷的基础。我们的结果揭示了神经元细胞命运决定因素在突触模式形成中通过抑制规范Wnt信号通路的新颖新颖的作用。


是激活的蛋白质促进BRAF V600E突变体转移性结直肠癌细胞对有丝分裂原激活的蛋白激酶途径
结直肠癌(CRC)在中国癌症中的发病率最高和第三高死亡率(1,2),大约15%的转移性CRC患者患有BRAF基因突变和预后不良(3,4)。Braf Oncogenes通过激活有丝分裂原活化蛋白激酶(MAPK)途径(5)来促进肿瘤发生,而V600E突变是最常见的BRAF突变。V600E突变患者的死亡风险是野生型BRAF患者的两倍,因为前者中的大多数人都对化疗和靶向药物疗法具有抗性。尽管一些小样本研究表明,与贝伐单抗相结合的三药化疗(FOLFOXIRI)可能会在某种程度上改善这些患者的预后,但生存益处似乎有限(6,7)。因此,对BRAF V600E突变的新型药物和治疗方案的探索构成了紧急的临床优先事项。
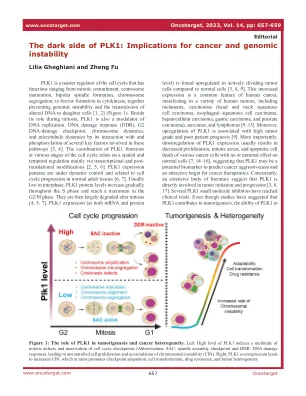
PLK1的阴暗面:对癌症和基因组不稳定性的影响
plk1是细胞周期的主要调节剂,其功能范围从有丝分裂承诺,中心体成熟,双极纺锤体形成,染色体分离,染色体分离,在细胞因子中的毛茸茸形成,共同防止基因组不稳定性和可预防基因组不稳定性和对女子细胞的传播到子细胞[1,2](图1)。在其在有丝分裂过程中的作用外,PLK1还是DNA复制,DNA损伤响应(DDR),G2 DNA损伤检查点,染色体动力学和微管动力学的调节剂,其与这些途径中涉及的几个关键因素的相互作用和磷酸化相互作用[3,4]。PLK1在细胞周期的各个阶段的协调依赖于空间和时间调节,主要是通过转录和翻译后修饰[2,5,6]。PLK1表达模式受到动态控制,并且与正常成人组织的细胞周期进程有关[6,7]。通常在相间的相间较低,PLK1蛋白水平在整个S相逐渐增加,并在G2/m相中达到最大值。然后,它们在有丝分裂后大大降解[4,5,7]。plk1表达(在mRNA和蛋白质上

在 PRC1 和 EB1 基因编辑细胞中,中央纺锤体的逐渐压缩降低了其动态性
在有丝分裂过程中,纺锤体会发生形态和动态变化。它在后期开始时重组,此时反平行束 PRC1 积累并将中央纺锤体蛋白募集到中间区。人们对中央纺锤体在人类细胞中形态变化过程中的动态特性如何变化知之甚少。利用基因编辑,我们生成了从其内源性荧光位点表达 PRC1 和 EB1 的人类细胞,以量化其天然纺锤体分布和结合/解离周转。EB1 正末端追踪显示微管生长普遍减慢,而 PRC1 与其酵母直系同源物 Ase1 类似,与压缩的反平行微管重叠结合越来越强。 KIF4A 和 CLASP1 与中央纺锤体的结合更具动态性,但也显示出减慢的周转速度。这些结果表明,中央纺锤体在有丝分裂过程中逐渐变得更加稳定,这与最近在有丝分裂后期中央纺锤体中反向平行中区束形成的“捆绑、滑动和压缩”模型一致。


核进口缺陷驱动神经退行性中的细胞周期失调
神经退行性疾病(NDDS)和其他与年龄有关的疾病已通过一组关键的病理标志在经典上定义。这些标志中的两个,细胞周期失调(CCD)和核质转运(NCT)缺陷,长期以来一直在争论为因果关系,在加速衰老的病理学中是因果关系。具体而言,已证明有丝分裂后神经元中异常细胞周期活化会触发神经元细胞死亡途径和细胞衰老。此外,已经观察到NCT在衰老和神经变性过程中逐渐失调,其中增加了核蛋白的亚细胞再分配(例如TAR DNA-结合蛋白43(TDP43))对细胞质的主要驱动力是许多NDDS的主要驱动力。然而,NCT缺陷的功能意义是作为病理学的主要驱动因素或后果,以及细胞周期机械的重新分布如何促进神经变性,尚不清楚。在这里,我们描述了对进口素β进口的药理抑制能够在丝分裂神经元细胞系和有丝分裂后原发性神经元体外扰动细胞周期机制。以核进口缺陷为特征的运动神经元疾病的NEMF R86S小鼠模型,进一步概括了有丝分裂细胞系中CCD的标志,在体外和有丝分裂后的原发性神经元中以及体内脊柱运动神经元中。观察到的CCD与NDDS中神经元细胞死亡和细胞衰老中观察到的转录和表型失调一致。在一起,这些证据表明,导致CCD的核进口途径受损可能是神经变性中病理学的常见驱动力。