XiaoMi-AI文件搜索系统
World File Search System着丝粒

植物着丝粒的结构、功能和进化
着丝粒是真核染色体的重要区域,负责形成着丝粒复合体,在细胞分裂过程中与纺锤体微管连接。值得注意的是,尽管着丝粒在染色体分离中保持保守功能,但其底层 DNA 序列在物种内和物种间都存在差异,并且主要是重复性的。着丝粒的重复内容包括高拷贝串联重复序列(卫星)和/或特定的转座子家族。着丝粒的功能区域由特定组蛋白 3 变体 (CENH3) 的加载定义,该变体使着丝粒成核并显示动态调节。在许多植物中,着丝粒由卫星重复阵列组成,这些阵列的 DNA 甲基化程度高,并被嗜着丝粒的逆转录转座子侵入。在某些情况下,逆转录转座子成为 CENH3 加载的位点。我们回顾了植物着丝粒的结构,包括单着丝粒、全着丝粒和元多着丝粒结构,这些结构在染色体上着丝粒附着位点的数量和分布上有所不同。我们讨论了 CENH3 负荷的变化如何在植物胚胎发生早期细胞分裂过程中驱动基因组消除。我们回顾了表观遗传状态如何影响着丝粒身份,并讨论了试图解释跨物种观察到的着丝粒序列快速变化的进化模型,包括重组的潜在作用。我们概述了可能在着丝粒内起作用的假定选择模式,以及重复序列在驱动着丝粒进化周期中的作用。虽然我们的主要重点是植物基因组,但我们将其与动物和真菌着丝粒进行了比较,以得出着丝粒结构和功能的真核生物范围的视角。

植物着丝粒的结构、功能和进化
着丝粒是真核染色体的重要区域,负责形成着丝粒复合体,在细胞分裂过程中与纺锤体微管连接。值得注意的是,尽管着丝粒在染色体分离中保持保守功能,但其底层 DNA 序列在物种内和物种间都存在差异,并且主要是重复性的。着丝粒的重复内容包括高拷贝串联重复序列(卫星)和/或特定的转座子家族。着丝粒的功能区域由特定组蛋白 3 变体 (CENH3) 的加载定义,该变体使着丝粒成核并显示动态调节。在许多植物中,着丝粒由卫星重复阵列组成,这些阵列的 DNA 甲基化程度高,并被嗜着丝粒的逆转录转座子侵入。在某些情况下,逆转录转座子成为 CENH3 加载的位点。我们回顾了植物着丝粒的结构,包括单着丝粒、全着丝粒和元多着丝粒结构,这些结构在染色体上着丝粒附着位点的数量和分布上有所不同。我们讨论了 CENH3 负荷的变化如何在植物胚胎发生早期细胞分裂过程中驱动基因组消除。我们回顾了表观遗传状态如何影响着丝粒身份,并讨论了试图解释跨物种观察到的着丝粒序列快速变化的进化模型,包括重组的潜在作用。我们概述了可能在着丝粒内起作用的假定选择模式,以及重复序列在驱动着丝粒进化周期中的作用。虽然我们的主要重点是植物基因组,但我们将其与动物和真菌着丝粒进行了比较,以得出着丝粒结构和功能的真核生物范围的视角。

着丝粒转座因子在塑造染色体进化中的作用
(> 11 百万年前),其特点是 Athila 和 CRM 元素贡献相等(模式:分别为 467 和 353 TE)。这些发现表明这些物种的着丝粒周围相对稳定,较旧的 CRM 副本随后被 Athila 元素所取代。相比之下,B. prealpina 和 B. varia 显示出更高的 CRM 序列周转率,许多旧的 CRM 副本被较新的副本所取代。在分布分散的 Athila 家族中也观察到了类似的模式。最后,我们扩展了

酵母和灵长类动物着丝粒 DNA 的隐藏结构特征
白质核酸酵素, 44, 1665 (1999). 3) L. Vernis, A. Abbas, M. Chasles, CM Gaillardin, C. Brun, JA Huber-

致病酵母近平滑念珠菌中多态性着丝粒的位置
着丝粒提出了一个进化悖论:功能高度保守,但序列和结构却迅速变化。然而,在没有损伤的情况下,着丝粒的位置通常在一个物种内是保守的。我们在此报告,致病酵母菌种近平滑假丝酵母的分离株在其八条染色体中的两条染色体上表现出着丝粒位置的种内多态性。它的旧着丝粒具有反向重复 (IR) 结构,而其新着丝粒没有明显的结构特征,但位于旧位置的 30 kb 以内。因此,着丝粒可以自然地从一个染色体位置移动到另一个染色体位置,似乎是自发的,并且在 DNA 序列没有任何显著变化的情况下。我们的观察结果与所有着丝粒都是由基因决定的模型相一致,例如由短或长 IR 的存在或形成十字形的能力决定。我们还发现着丝粒已成为 C. parapsilosis 进化枝中基因组重排的热点。
间充质肿瘤和白血病中着丝粒聚集的高患病率和依赖性
多余着丝粒的存在是癌症的一个标志,在侵袭性肿瘤中经常观察到。着丝粒扩增的癌细胞通过特定的应对机制实现伪双极纺锤体以求生存。然而,它们在癌症中的分布和流行程度仍然很大程度上未知。在这里,我们使用 NCI60 组癌细胞系,表明应对策略的存在与着丝粒扩增相关,其中两个纺锤体极内额外着丝粒的聚集是最普遍的机制。此外,我们报告了着丝粒聚集能力与上皮-间质转化 (EMT) 之间的关联,并观察到着丝粒扩增的乳腺癌细胞中间质特征的诱导促进了聚集。

研究文章 CUT&RUN 识别了 Rhodotorula toruloides IFO0880 的着丝粒 DNA 区域
人们越来越多地研究将红酵母用作脂质、脂肪酸衍生物和萜类化合物的生物生产宿主。人们已经开发了各种遗传工具,但尚未报道过着丝粒和自主复制序列 (ARS),而这两者都是维持稳定的游离质粒所必需的元素。在本研究中,使用靶标下切割并使用核酸酶释放 (CUT&RUN)(一种用于全基因组 DNA-蛋白质相互作用映射的方法)来识别与着丝粒组蛋白 H3 蛋白 Cse4(着丝粒 DNA 的标记)相关的红酵母 IFO0880 基因组区域。识别并分析了 15 个长度从 8 到 19 kb 不等的假定着丝粒,并对其中四个进行了 ARS 活性测试,但未显示 ARS 活性。这些着丝粒序列含有低于平均水平的 GC 含量,对应于转录冷点,主要是非重复的,并且共享一些残留转座子相关序列,但除此之外没有显示显著的序列保守性。未来在该酵母中识别 ARS 的努力可以利用这些着丝粒 DNA 序列来提高来自假定 ARS 元素的游离质粒的稳定性。

对诺斯卡品化合物作为靶向微管蛋白受体的抗肿瘤药物进行计算机分析
科学研究表明,微管蛋白在人体不同部位的肿瘤中都有表达。III 类 b 型微管蛋白 (TUB b 3 ) 是与晚期肿瘤相关的最主要的微管蛋白。38 蛋白质研究表明,微管蛋白在细胞行为中起着至关重要的作用,是微管的结构单位。着丝粒可以作为癌症进展的指标进行监测;这种结构是一种动态元素,可组织负责细胞分裂的机制。着丝粒由一对中心粒组成,纺锤体和星状微管由此起源。癌细胞通常具有额外的着丝粒和染色体不稳定性。着丝粒的这些数值和结构变化是人类癌症和染色体疾病的有用指标和标志。人体组织含有各种
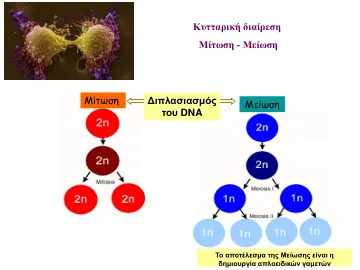

TD 2(DNA复制和特性传递)
- 在 ................................. 前期,核膜碎裂成碎片 - 在 ................................. 中期,纺锤体有丝分裂的赤道板形成 - 在 ................................. 中期,染色单体分离形成两组子染色体 - DNA 合成的时期称为 S 期 - 纺锤体有丝分裂由微管组成,微管是亚基微管蛋白的聚合物 - 染色体迁移是通过纺锤体微管与与每个染色体的着丝粒相关的结构结合实现的:着丝粒