XiaoMi-AI文件搜索系统
World File Search System端缝

继发于钩端螺旋体病的完整心脏块
抽象的完全心脏阻滞(CHB)是一种罕见但潜在的威胁生命的并发症,这是由小卵磷酸属引起的人畜共患细菌感染。虽然钩端螺旋体病主要影响肾脏和肝脏,但包括CHB在内的心脏受累可能会发生并具有显着的临床意义。钩端螺旋体病中CHB的发病机理是多因素的,可能涉及钩端螺旋体生物,全身性炎症反应,自身免疫反应,电解质失衡和血液动力学作用的直接心脏侵袭。迅速识别和对CHB的管理对于防止不良后果,包括血液动力学不稳定和心脏猝死至关重要。治疗策略包括诸如血液动力学支持和电解质失衡的纠正,有症状性心动过缓的临时起搏,基础感染的抗生素疗法以及在折磨情况下考虑永久起搏器植入的抗生素治疗。。心脏表现可能包括心肌炎,心包炎,心律不齐,传导阻滞和心脏衰竭。我们报告了一例钩端螺旋体病在以前健康的年轻绅士中引起心脏障碍。

复合材料行业供应商 - 硬质塑料
代码 描述 尺寸 FGTISS/M2 表面组织 FGCMxx/KG 短切纤维垫 300 和 450gm/m2 FGWRxx/KG 编织粗纱(单向和双向) 450 和 600 gm/m2 缝织物 FG-缝织物 各种缝织物 布 FGCSPI/xx 平纹玻璃布 140、200、300gm/m2 FGCTWL-xx 斜纹玻璃布 200 gm/m2 CFC-200PL/M2 碳纤维平纹 200 gm/m2 CFC-200TWL/M2 碳纤维斜纹 200 gm/m2 注意 +-140gm/m2 = 4 oz, +-200gm/m2 = 6oz, +-300gm/m2 = 10盎司布

关于量子隧穿时间的现状 | PhilSci-Archive
我们有一种天真的古典直觉,认为我们最好的理论应该能够告诉我们物理过程的持续时间。受这种简单的古典图景的启发,物理学家们问道,量子粒子穿过经典禁能垒需要多长时间?换句话说,量子隧穿时间的正确表达式是什么?与经典问题不同,这个问题似乎没有一个直接的答案,并在物理学文献中引发了广泛的争论。物理学家提出了各种量子隧穿时间的表达式。一些跟踪隧穿系统的内部特性,而另一些则依赖于隧穿粒子和外部物理系统之间的耦合。一般来说,它们都提供了不同的值——只在某些限制内相一致——并且它们在大多实用的基础上相互权衡。然而,一些作者仍然在谈论,好像有一个明确而独特的表达可以找到,或者至少好像一些提出的表达本质上比其他表达更有意义。许多人认为,这种明显的歧义源于量子力学对待时间的一般方式:将其视为参数,而非算符。其他人则强调了这场争论的解释维度,甚至认为隧穿时间在量子力学的标准解释中毫无意义。然而,这种混乱和歧义只存在于标准的“正统”或“哥本哈根”解释中——所有考虑德布罗意-玻姆“导波”解释传统形式的作者都同意,这种解释为隧穿时间提供了一个清晰明确的表达,其中量子态由受波函数演化引导的物理德布罗意-玻姆粒子组成。这引发了人们的猜测:量子隧穿时间的实验测试是否可以作为传统形式的德布罗意-玻姆理论的实验测试。因此,关于量子隧穿时间的文献现状自然而然地引出了三个物理和哲学问题。首先,关于隧穿时间的困惑是否真的源于量子力学中更普遍的“时间问题”——即时间缺乏算符这一事实?其次,隧穿时间在量子力学的标准解释中真的是一个毫无意义的概念吗?如果是,为什么?最后,原则上,是否可以使用量子隧穿时间的实验测试作为德布罗意-玻姆解释的实验测试?本文旨在依次回答每个问题。自始至终,我都局限于德布罗意-玻姆理论的传统版本,其中隧穿时间是清晰明确的——其他关于导航波程序所依据的本体论的提议,虽然本身就很吸引人,但与我要提出的概念点无关。在本文的前半部分,即第 2 节中,我概述了现有的关于量子隧穿时间的文献。第 2.1 节解释了隧穿时间讨论所基于的物理场景。在第 2.2 节中,我描述了时间在量子力学中的一些特征,并展示了这些特征是如何被用来将量子隧穿时间的混乱归咎于量子力学中更普遍的“时间问题”。在第 2.3 节中,我描述了隧穿时间与量子力学解释之间的联系,并展示了这种联系是如何被用来激发两种主张的:关于标准解释中隧穿时间的意义的主张,以及关于使用隧穿时间作为 Bohmian 计划的“关键”实验测试的可能性的主张。在本文的后半部分,即第 3 节,我提出了自己的分析,为上述三个问题的答案辩护。我首先在隧穿问题和众所周知的双缝实验之间建立了一个类比。我指出,尝试建立特定于传输粒子的隧穿时间类似于尝试确定检测到的粒子是通过双缝的左缝还是右缝(第 3.1 节)。这个简单而有力的类比将构成本文其余部分的概念基础。接下来,将在第 3.2、3.3 和 3.4 节中回答这三个问题。至于围绕量子隧穿时间的明显混乱和模糊性是否可以追溯到量子力学中更普遍的时间问题,我认为“不”:混乱的真正根源是叠加,因此,即使时间可以用算符表示,隧穿时间在量子力学的标准解释中也是模糊和有争议的(第 3.2 节)。至于隧穿时间在量子力学的标准解释中是否毫无意义:我认为它与询问粒子是通过双缝实验的左缝还是右缝一样毫无意义(第 3.3 节)。最后,关于是否可能在原则上将量子隧穿时间用作德布罗意-玻姆解释的实验测试:我旨在提供一个简单的解释,说明为什么这是不可能的。不可能通过实验测量德布罗意-玻姆理论预测的隧穿时间,就像不可能测量粒子是通过左缝还是右缝而使屏幕上的干涉图案保持完整(第 3.4 节)一样。这些答案并不全是新的。文献中已经暗示了每一个,但它们尚未联系在一起——当它们出现时,它们被插入更长的简短评论中我描述了时间在量子力学中的一些特征,并展示了这些特征是如何被用来将量子隧穿时间的混乱归咎于量子力学中更普遍的“时间问题”。在第 2.3 节中,我描述了隧穿时间与量子力学解释之间的联系,并展示了这种联系是如何被用来激发两种主张的:关于隧穿时间在标准解释中的意义的主张,以及关于使用隧穿时间作为 Bohmian 计划的“关键”实验测试的可能性的主张。在本文的后半部分,即第 3 节,我提出了自己的分析,为上述三个问题的答案辩护。我首先在隧穿问题和众所周知的双缝实验之间建立了一个类比。我表明,尝试建立特定于透射粒子的隧穿时间类似于尝试确定检测到的粒子是通过双缝的左侧通道还是右侧通道(第 3.1 节)。这个简单而有力的类比将构成本文其余部分的概念基础。接下来,我们将在第 3.2、3.3 和 3.4 节中回答这三个问题。至于围绕量子隧穿时间的明显混乱和模糊性是否可以追溯到量子力学中更普遍的时间问题,我认为“不能”:混乱的真正根源是叠加,因此,即使时间可以用算符表示,隧穿时间在量子力学的标准解释中也是模糊和有争议的(第 3.2 节)。至于隧穿时间在量子力学的标准解释中是否毫无意义:我认为它的意义不亚于询问粒子是通过双缝实验的左缝还是右缝(第 3.3 节)。最后,至于原则上是否可以将量子隧穿时间用作德布罗意-玻姆解释的实验测试:我旨在提供一个简单的解释,说明为什么这是不可能的。不可能通过实验测量德布罗意-玻姆理论预测的隧穿时间,就像不可能测量粒子是通过左缝还是右缝而使屏幕上的干涉图样保持完整(第 3.4 节)一样。这些答案并不全是新的。文献中已经暗示了每一个,但它们尚未联系在一起——而且它们确实出现的地方,都是作为简短的评论插入到更长的我描述了时间在量子力学中的一些特征,并展示了这些特征是如何被用来将量子隧穿时间的混乱归咎于量子力学中更普遍的“时间问题”。在第 2.3 节中,我描述了隧穿时间与量子力学解释之间的联系,并展示了这种联系是如何被用来激发两种主张的:关于隧穿时间在标准解释中的意义的主张,以及关于使用隧穿时间作为 Bohmian 计划的“关键”实验测试的可能性的主张。在本文的后半部分,即第 3 节,我提出了自己的分析,为上述三个问题的答案辩护。我首先在隧穿问题和众所周知的双缝实验之间建立了一个类比。我表明,尝试建立特定于透射粒子的隧穿时间类似于尝试确定检测到的粒子是通过双缝的左侧通道还是右侧通道(第 3.1 节)。这个简单而有力的类比将构成本文其余部分的概念基础。接下来,我们将在第 3.2、3.3 和 3.4 节中回答这三个问题。至于围绕量子隧穿时间的明显混乱和模糊性是否可以追溯到量子力学中更普遍的时间问题,我认为“不能”:混乱的真正根源是叠加,因此,即使时间可以用算符表示,隧穿时间在量子力学的标准解释中也是模糊和有争议的(第 3.2 节)。至于隧穿时间在量子力学的标准解释中是否毫无意义:我认为它的意义不亚于询问粒子是通过双缝实验的左缝还是右缝(第 3.3 节)。最后,至于原则上是否可以将量子隧穿时间用作德布罗意-玻姆解释的实验测试:我旨在提供一个简单的解释,说明为什么这是不可能的。不可能通过实验测量德布罗意-玻姆理论预测的隧穿时间,就像不可能测量粒子是通过左缝还是右缝而使屏幕上的干涉图样保持完整(第 3.4 节)一样。这些答案并不全是新的。文献中已经暗示了每一个,但它们尚未联系在一起——而且它们确实出现的地方,都是作为简短的评论插入到更长的并声称可以使用隧穿时间作为 Bohmian 方案的“关键”实验测试。在本文的后半部分,即第 3 节中,我将提出自己的分析,为上述三个问题提供答案。我首先在隧穿问题和众所周知的双缝实验之间建立类比。我表明,尝试建立特定于透射粒子的隧穿时间类似于尝试确定检测到的粒子是通过双缝的左侧通道还是右侧通道(第 3.1 节)。这个简单而有力的类比将构成本文其余部分的概念基础。然后在第 3.2、3.3 和 3.4 节中回答这三个问题。至于围绕量子隧穿时间的明显混乱和模糊性是否可以追溯到量子力学中更普遍的时间问题,我认为“不能”:混乱的真正根源是叠加,因此,即使时间可以用算符表示,隧穿时间在量子力学的标准解释中也是模糊和有争议的(第 3.2 节)。至于隧穿时间在量子力学的标准解释中是否毫无意义:我认为它与询问粒子是否穿过双缝实验的左缝或右缝一样毫无意义(第 3.3 节)。最后,至于原则上是否可以将量子隧穿时间用作德布罗意-玻姆解释的实验测试:我旨在提供一个简单的解释,说明为什么这是不可能的。不可能通过实验测量德布罗意-玻姆理论预测的隧穿时间,就像不可能测量粒子是通过左缝还是右缝而使屏幕上的干涉图样保持完整(第 3.4 节)一样。这些答案并不全是新的。文献中已经暗示了每一个,但它们尚未联系在一起——而且它们确实出现的地方,都是作为简短的评论插入到更长的并声称可以使用隧穿时间作为 Bohmian 方案的“关键”实验测试。在本文的后半部分,即第 3 节中,我将提出自己的分析,为上述三个问题提供答案。我首先在隧穿问题和众所周知的双缝实验之间建立类比。我表明,尝试建立特定于透射粒子的隧穿时间类似于尝试确定检测到的粒子是通过双缝的左侧通道还是右侧通道(第 3.1 节)。这个简单而有力的类比将构成本文其余部分的概念基础。然后在第 3.2、3.3 和 3.4 节中回答这三个问题。至于围绕量子隧穿时间的明显混乱和模糊性是否可以追溯到量子力学中更普遍的时间问题,我认为“不能”:混乱的真正根源是叠加,因此,即使时间可以用算符表示,隧穿时间在量子力学的标准解释中也是模糊和有争议的(第 3.2 节)。至于隧穿时间在量子力学的标准解释中是否毫无意义:我认为它与询问粒子是否穿过双缝实验的左缝或右缝一样毫无意义(第 3.3 节)。最后,至于原则上是否可以将量子隧穿时间用作德布罗意-玻姆解释的实验测试:我旨在提供一个简单的解释,说明为什么这是不可能的。不可能通过实验测量德布罗意-玻姆理论预测的隧穿时间,就像不可能测量粒子是通过左缝还是右缝而使屏幕上的干涉图样保持完整(第 3.4 节)一样。这些答案并不全是新的。文献中已经暗示了每一个,但它们尚未联系在一起——而且它们确实出现的地方,都是作为简短的评论插入到更长的我认为“不”:真正的混乱根源是叠加,因此即使时间可以用算符表示,隧穿时间在量子力学的标准解释中也是含糊不清且有争议的(第 3.2 节)。至于隧穿时间在量子力学的标准解释中是否毫无意义:我认为它与询问粒子是通过双缝实验的左缝还是右缝一样毫无意义(第 3.3 节)。最后,至于原则上是否可以将量子隧穿时间用作德布罗意-玻姆解释的实验测试:我旨在提供一个简单的解释,说明为什么这是不可能的。实验测量德布罗意-玻姆理论预测的隧穿时间是不可能的,就像测量粒子是通过左缝还是右缝而不使屏幕上的干涉图案保持完整一样(第 3.4 节)。这些答案并不都是新的。文献中已经提到过每一个,但它们还没有联系在一起——即使它们出现了,它们也会作为简短的评论插入到更长的我认为“不”:真正的混乱根源是叠加,因此即使时间可以用算符表示,隧穿时间在量子力学的标准解释中也是含糊不清且有争议的(第 3.2 节)。至于隧穿时间在量子力学的标准解释中是否毫无意义:我认为它与询问粒子是通过双缝实验的左缝还是右缝一样毫无意义(第 3.3 节)。最后,至于原则上是否可以将量子隧穿时间用作德布罗意-玻姆解释的实验测试:我旨在提供一个简单的解释,说明为什么这是不可能的。实验测量德布罗意-玻姆理论预测的隧穿时间是不可能的,就像测量粒子是通过左缝还是右缝而不使屏幕上的干涉图案保持完整一样(第 3.4 节)。这些答案并不都是新的。文献中已经提到过每一个,但它们还没有联系在一起——即使它们出现了,它们也会作为简短的评论插入到更长的

富满微电子集团股份有限公司
供电电源 VDD ........................................................................................................................................... .. -0.3V~+10V VM 、 COUT 端允许输入电压 .................................................................................................. ....VDD-25V~VDD+0.3V DOUT 端允许输入电压 ......................................................................................................................- 0.3V~VDD+0.3V 工作温度 TA ..................................................................................................................................................- 40 ℃ ~+85 ℃ 结温 ........................................................................................................................................................................... 150 ℃ 贮存温度 .......................................................................................................................................................- 65 ℃ ~150 ℃ 功耗 PD ( TA=25 ℃) SOT23-6 封装(热阻 θJA = 200 ℃ /W ) .................................................................. ..625mW 焊接温度(锡焊, 10 秒) ..................................................................................................................................... 260 ℃

在西里兰卡西部省份的钩端螺旋体病患者样本中检测到的钩端螺旋体探究的新型FLAB基因变异
已描述了四种flab1,flab1,flab2,flab3和flab4)的同种型[35]。,大多数研究都将FLAB2基因作为其靶标[26,28]。钩端螺旋体的鞭毛细丝,显示一个复杂的结构,该结构由由鞘蛋白(281至285个氨基酸)制成的中心核心组成,周围环绕

Eurican L4,INN-犬钩端螺旋体病疫苗(灭活)
1 小于 6 cm,8 天内消失 2 2 天内消失 3 3 天内消失 4 4 天内消失 5 最高 39.8 °C,1 天内消失。 6 包括可能危及生命的过敏性休克。如果发生此类反应,应立即采取适当的治疗措施。报告不良事件很重要。它允许对兽药进行持续的安全监测。报告应通过兽医发送给上市许可持有人或其当地代表或通过国家报告系统发送给国家主管当局,最好通过兽医。请参阅包装说明书的“联系方式”部分。 3.7 在怀孕、哺乳或产仔期间使用 勃林格殷格翰三价钩端螺旋体疫苗(含犬钩端螺旋体、黄疸出血钩端螺旋体和流感伤寒钩端螺旋体)的怀孕母犬的安全数据表明,该疫苗可在怀孕期间使用。对于含有额外灭活菌株澳大利亚钩端螺旋体的 Eurican L4,尚无关于怀孕母犬的安全数据。 3.8 与其他药物的相互作用和其他形式的相互作用 有安全性和有效性数据表明,该疫苗可与 Eurican DAP 或 Eurican DAPPi/Eurican DHPPi 混合使用。有安全性和有效性数据表明,对于 12 周龄以上的犬,该疫苗可以与 Rabisin 同一天接种,但不能与 Rabisin 混合。除了上述产品外,没有关于此疫苗与任何其他兽药一起使用时的安全性和有效性的信息。因此,需要根据具体情况决定在任何其他兽药之前或之后使用此疫苗。3.9 给药途径和剂量当单独使用 Eurican L4 时,皮下注射 1 毫升剂量。当 Eurican L4 用作 Eurican DAP 或 Eurican DAPPi / Eurican DHPPi 的稀释剂时,用 Eurican L4 疫苗悬浮液无菌重构冻干物的内容物。使用前充分混合。重构小瓶的全部内容物应作为单剂量给药。

近端小管缩短对Lowe综合征模型中蛋白质排泄的影响
底物是内吞作用的主要调节剂,预计LS LS患者的LMW蛋白尿是由于PT顶端内吞途径沿PT的某些有效功能所致。3与此相一致,培养细胞模型中的一部分研究表明,OCRL在内吞回收中起作用,这是通过防止在内吞囊泡上积累的肌动蛋白涂层的解聚和/或回收箱的作用。4,5但是,OCRL在细胞稳态中也具有许多其他角色,包括睫状生物发生,6-8细胞极性和自噬。6,9,10此外,OCRL在细胞因子期间被招募到脱落部位。11 ptdins(4,5)p 2累积稳定在细胞因子过程中的细胞内桥,并且其通过OCRL的水解对于脱落是必要的。11尚不清楚这些功能如何促进LS病理学。另一个未解决的问题是,OCRL的损失如何损害LS患者的Ca 2+,HCO 3 2和氨基酸的PT恢复。近年来已经开发了LS的小鼠和斑马鱼模型,但是在细胞培养中观察到的分子和细胞缺陷与患者和动物模型的表型之间的联系仍然难以捉摸。缺乏OCRL的转基因斑马鱼表现出降低的巨蛋白水平,降低了流体相位标记物的上升水平,除了与LS患者观察到的患者一致的眼睛和面部缺陷外,促脑肾脏PT中的亚皮囊泡较少。8,12小鼠LS模型的开发更为复杂。 这些8,12小鼠LS模型的开发更为复杂。这些OCRL敲除(KO)小鼠没有明显的表型,因为它们表达了高水平的Inpp5b,这是另一种磷脂酰肌醇5 9-磷酸酶,显然可以对某些OCRL功能进行操作。13 - 15小鼠PT中的inpp5b在较高水平和与人类相比的剪接变体中表达不同。16由于小鼠中的OCRL和INPP5B的全局KO是致命的,因此通过跨越OCRL KO小鼠的OCRL KO小鼠产生了17,18 LS小鼠模型,该小鼠过表达人Inpp5b与小鼠INPP5B KO:由此产生的雄性小鼠在年龄的8周时表现出适中的蛋白尿和氨基尿症。19,20已描述了一个最近的小鼠模型,其中在OCRL KO小鼠的肾脏中有条件地灭活了INPP5B。这些小鼠中的21个PT细胞表达了巨蛋白水平降低,并且表现出严重受损的内吞作用。令人惊讶的是,在KO之后没有立即观察到蛋白尿,而是需要几个月的发展。此时间滞后与OCRL对内吞途径功能的直接影响不一致,并表明在更长的时间段内发生的其他变化与LS表型相关。此外,需要靶向OCRL和INPP5B以观察任何肾脏表型,这是努力确定OCRL在Pt功能中的特定作用的努力。为了研究OCRL的损失如何影响PT功能,我们产生了PT细胞中LS的慢性CRISPR/CAS9 OCRL KO和LS的急性siRNA敲低模型。引人注目的是,在我们的所有模型中以及在患者纤维细胞中,我们观察到功能性OCRL的损失延长了细胞分裂的持续时间,并导致了多核细胞的积累。

umer aa。埃塞俄比亚钩端螺旋体病的一种健康影响
钩端螺旋体病是由诱发属的致病革兰氏阴性细菌引起的全球重要人畜共患病。该疾病发生在几乎所有哺乳动物的物种中,由于环境中的生存生物的较长,在热带地区更为常见。在受污染的环境中经常暴露于动物和人类中,表明一种健康方法。它是由属于钩端螺旋体属的许多血清中的血清引起的。钩端螺旋体引发并不是唯一类似于这种疾病的血清,几乎影响了所有哺乳动物。主要的储层动物被称为大鼠和小鼠。通过裸露的粘膜膜和皮肤受损的皮肤直接传播到感染动物的尿液中。对各种动物物种的实验室测试不会显着改变钩端螺旋体病的临床指征。该疾病的最佳控制是疫苗接种,隔离和啮齿动物管理。具有温暖,潮湿的天气和碱性或中性土壤的热带地区更适合钩端螺旋体生存。建议采用有效的控制措施并提高公众对钩端螺旋体病的自动传播的认识。相关的身体应参与支持埃塞俄比亚等被剥夺国家的钩端螺旋体病的研究。
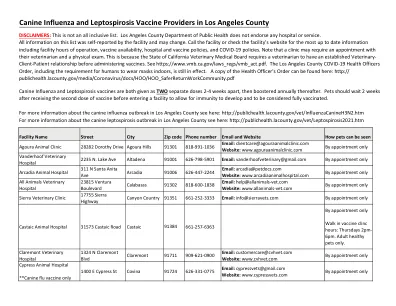
洛杉矶县犬类流感和钩端螺旋体病的提供者
免责声明:这不是全包列表。洛杉矶县公共卫生部不认可任何医院或服务。此列表上的所有信息都是由该设施自我报告的,可能会改变。致电该设施或检查设施的网站以获取最新信息,包括设施小时的操作,疫苗可用性,医院和疫苗策略以及COVID-19-19策略。请注意,诊所可能需要与兽医和身体检查一起任命。这是因为加利福尼亚州的兽医医疗委员会要求兽医在管理疫苗之前具有既定的兽医与客户关系。请参阅https://www.vmb.ca.gov/laws_regs/vmb_act.pdf。洛杉矶县共同卫生官员命令,包括人类在室内戴口罩的要求仍然有效。可以在此处找到卫生官员命令的副本:http://publichealth.lacounty.gov/media/coronavirus/docs/hoo/hoo/hoo_saferreturnworkcommunity.pdf
翠鸟柔性自动隔离病毒DNA/RNA的方案| neb
将打靶特定人源基因的 Cas9 和 sgRNA 转染到 HEK293 细胞。转染所用的质粒 DNA 上含有 表达带双端核定位序列 ( NLS )的 Cas9 及 sgRNA 的表达框,通过 TransIT-X2 (Mirus) 转染 试剂进行转染。转染所用的 Cas9 mRNA 进行了假尿苷和 5- 甲基胞嘧啶修饰且带有双端 核定位序列,使用 transIT-mRNA 转染试剂将 sgRNA 和 mRNA 共转染。 Cas9 RNPs 使用脂质 体 RNAiMAX ( Life Technologies ) 进行反向转染, RNP 的终浓度为 10 nmol 。 Cas9 蛋白上不含 核定位序列。 EnGen Cas9 含有双端核定位序列。编辑效率通过 T7E1 实验进行分析,结果 以修饰百分比进行统计。